
Abstract
Three tree species, Norway maple, green ash, and ginkgo were transplanted at four different periods over the course of the 1979 growing season. Root regeneration was measured 1 year after transplanting. Regenerated roots were initiated primarily from the severed root ends. Norway maple showed the greatest capacity for root regeneration during this period. The availability of carbohydrates to support this new root growth was limited only during the period of active shoot growth in the early spring. Transplanting trees during this period should be avoided.
Rapid regeneration of the root system is probably the most important factor for the successful establishment of transplanted trees and other woody species. Any factor that reduces the rate of root growth will decrease the probability of survival of transplants. Root initiation in transplanted trees occurs primarily near the end of the severed roots (1,15), particularly from starch-laden parenchyma cells in close proximity to vascular tissue (2,5).
Root growth is regulated by a number of physiological factors including the amounts of available carbohydrates. Any factor that affects the photosynthetic production of the shoot markedly affects root growth and elongation and may be a factor in the establishment of seedlings. Light, and its effect on photosynthesis, is an important environmental factor (14). Drought stress and defoliation also reduce photosynthesis and, therefore, root growth (9,10). Carbohydrates accumulate at the base of severed or girdled stem cuttings, and rooting can be enhanced if a shoot is girdled before the cutting is taken (13). It is not known whether the same phenomenon occurs at the ends of severed roots of transplanted trees.
In many deciduous trees, carbohydrate reserve levels normally decrease sharply during spring growth and reach a minimum in early summer. The carbohydrate levels subsequently increase to an autumn peak and finally decline slightly during the winter (6,12). The onset of spring growth is believed responsible for the sharp spring decrease in reserve carbohydrates.
The studies reported here were undertaken to characterize root regeneration patterns, to investigate the changes in carbohydrate reserves imposed by transplanting stresses, and to determine the effect of seasonal variation of carbohydrate reserves on root regeneration.
Methods
Transplanting
A total of 36 trees of three species, Norway maple, green ash, and ginkgo, was transplanted at four different times over the course of the 1979 growing season (Table 1). The trees, 5 to 10 cm in diameter, were moved in groups of four per species with a 44-inch (1,12-m) tree spade. The trees were not moved more than a few hundred yards. Consequently, disruption of the root ball was minimized, and the new soil was essentially the same type. Posttransplanting care involved limited pruning, regular watering, wrapping the trunk with standard tree wrap, and mulching the area around the base of each tree bounded roughly by the drip line.
Months in which three tree species were transplanted during the 1979 growing season.
Measurement of root regeneration after transplanting
A soil-core sampling method was used to measure regenerated roots on transplanted trees 1 year after the date of transplanting and on nontransplanted trees during June, 1980. The soil-core dimensions were 7.5 cm in diameter and 20 cm long. Each tree was sampled on four sides at the edge of the original root ball. On one randomly selected side of each tree, a second core was taken immediately below the first to a depth of 40 cm. This soil-sampling procedure was repeated in a second ring of four cores adjacent to the outside edge of the first cores. Each of the upper 20-cm samples was divided into equal upper and lower portions. For replicates of each tree species, samples of similar locations and depths were combined and treated as one sample. After removing all soil and roots of herbaceous plants and other woody species, total length measurements were taken on each set of root samples. Newman’s line-intersection method was used for estimating the total length of root in the sample. Length measurements were used to calculate root densities (meters of root length per liter of soil volume).
Carbohydrate assays
Tissue samples used for carbohydrate analysis were collected from twigs and roots 1 year after transplanting. Nontransplanted groups of similar tree species, sampled on the same date as the transplanted trees, served as controls. Samples were immediately returned to the laboratory and were stored at -20 C until analyzed.
The determination of food reserves (primarily nonstructural carbohydrates) was performed by using a modification of Priestley’s (11) analytical method. Four-gram frozen samples were cut into discs (bark and wood included), ground in a Virtis R-23 homogenizer for 1 minute, divided into four subsamples, dried at 100 C for 12 hours, and weighed to the nearest 0.1 mg. Each subsample was then wrapped in filter paper, tied with cotton thread, oven dried for 12 hours and weighed as above. Subsample extractions were taken by boiling each subsample for 2 hours in an aqueous solution of 5-percent trichloroacetic acid (TCA) and 35-percent methanol, followed by water extraction in a soxhlet apparatus for 5 hours. The wrapped subsamples were again oven dried and weighed. Dry weight loss, expressed as a percentage of original dry weight, was used as a measure of food reserves.
Results
Root regeneration after transplanting
Root densities of undisturbed control trees were higher than those of transplants of the same species for all seasons and at all depths. Absolute root density measurements showed that Norway maple consistently produced the greatest root growth and ginkgo produced the least growth of the three species (Fig. 1). However, the ginkgo control trees also had the least root density. If root regeneration is expressed as a percentage of the control, Norway maple again shows the highest rate of regeneration at 52 percent, while ginkgo and green ash show regeneration rates of 42 and 25 percent, respectively (Fig. 2).

Root regeneration of transplanted trees after 1 year of growth. (Stippled portions represent inner core samples taken 0 to 7.5 cm from the root ball edge. Slashes areas represent outer core samples taken 7.5 to 15 cm from the edge of the root ball. Control tree samples were taken at similar distances from the trunk, but were analyzed together.
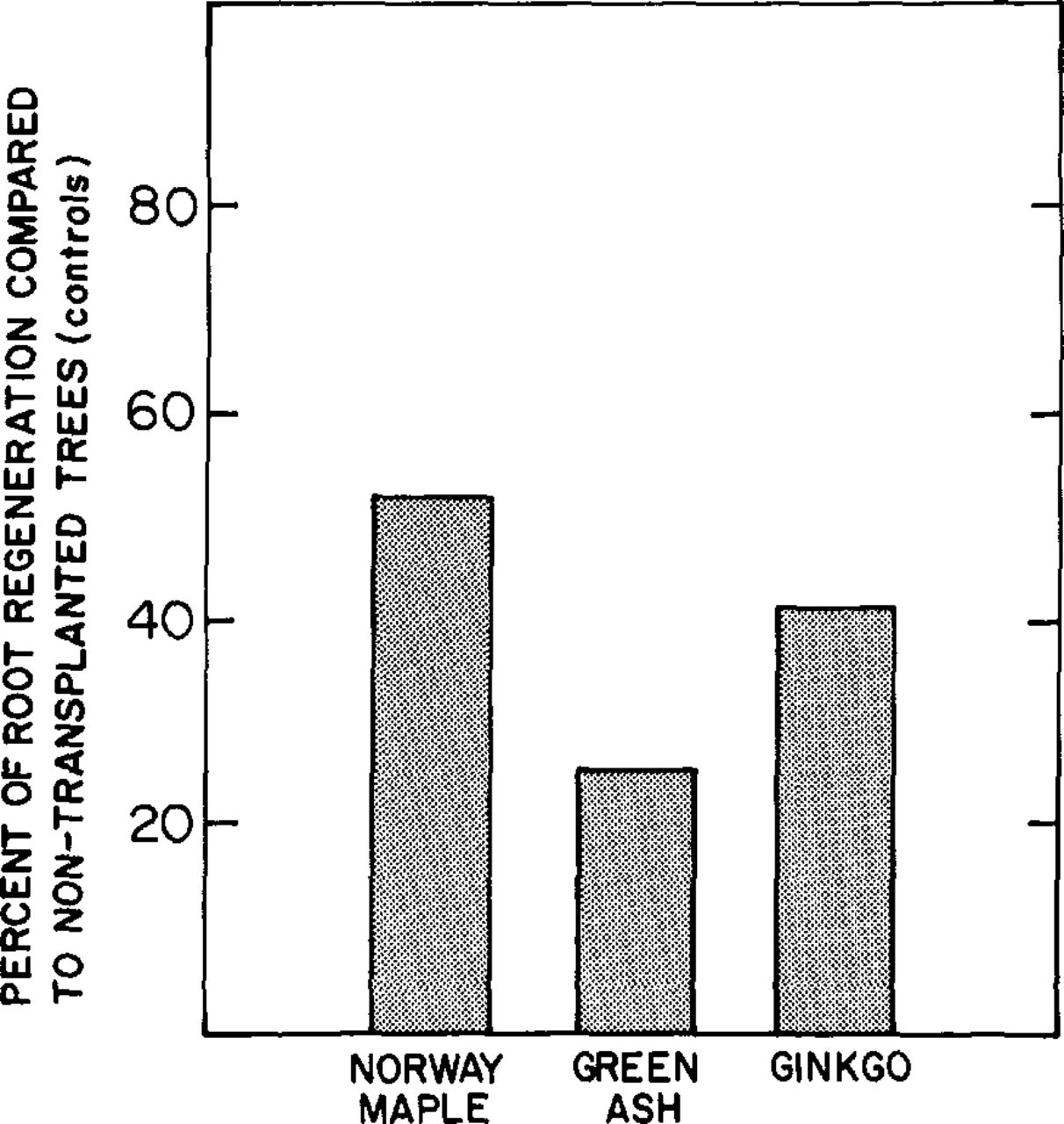
Relative root regeneration of transplanted trees in relation to fine-root densities of nontransplanted trees.
Season of transplanting appeared to influence root regeneration in Norway maples. Late spring (May) transplants showed the best root growth of all groups of Norway maples, and early spring (March) transplants showed the poorest growth. Green ash and ginkgo did not show significant variations in root regeneration levels with the transplant seasons.
For the transplanted trees, the vertical and horizontal distribution of fine roots within the coresampling pattern was similar to that of the controls. Root densities in the upper 20 cm of soil were as much as 33 percent higher than densities in the lower 20 cm. As expected, root densities were consistently higher in the soil closest to the original root balls of transplanted trees.
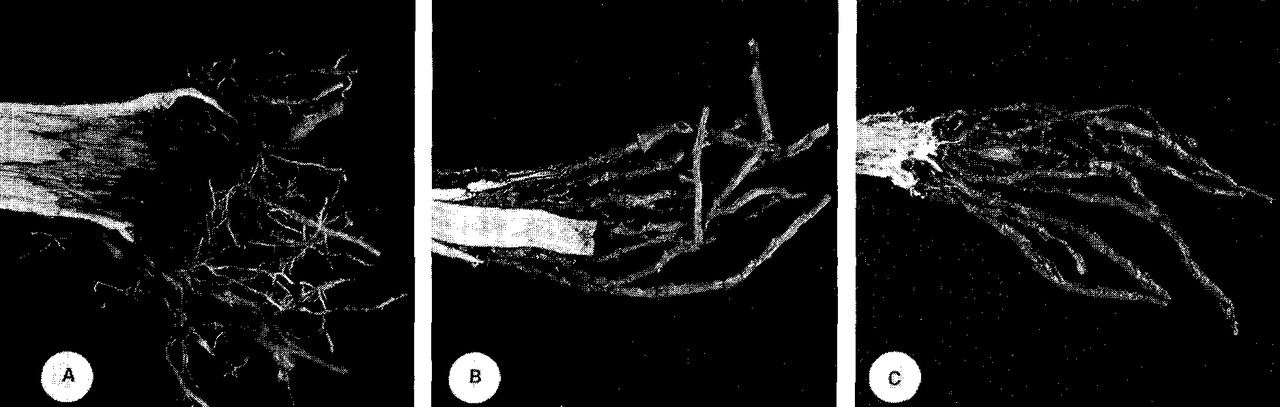
Regenerated roots originated almost exclusively from the callus tissue formed around the outer margins of the roots that were severed during transplanting (Fig. 3). The amount of small-root regeneration at the edge of the ball varied somewhat with species, but in general, few small roots originated from the callus zone of large roots (4 to 8 cm in diameter). Smaller roots (less than 4 cm in diameter) produced many new roots. On those species studied, significant amounts of new root development occurred only near the severed root ends at the outside edge of the root ball.

Regenerated roots originating from the callus tissue formed as a result of severing, (a) Norway maple root showing discoloration of the wood formed prior to transplanting, (b) Ginkgo root showing dieback of the cambium prior to root regeneration, (c) Same ginkgo root as (b) with bark removed, showing the locations of root initiation.
Carbohydrate assays
Assays of root and twig samples of nontransplanted Norway maple and green ash trees showed typical seasonal fluctuation of total carbohydrates (minimum in spring, maximum in fall) (Fig. 4). The assayed samples contained both bark and wood. Roots consistently had higher carbohydrate levels than corresponding twigs had. The assayed samples from the transplanted trees showed seasonal curves with steady or decreasing carbohydrate levels from July to October. Root tissue from nontransplanted trees showed an increase in total carbohydrates during this period. The carbohydrate levels of transplanted trees were higher than those of controls during the spring and summer, and were slightly lower than the levels of controls in the fall.
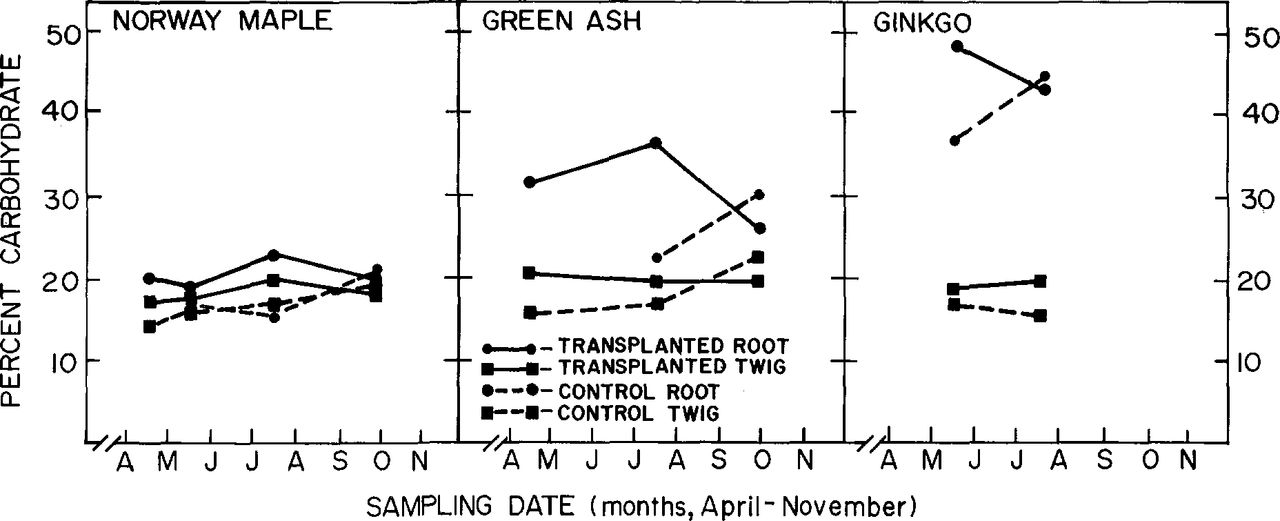
Total carbohydrate content in root wood and bark tissue of transplanted and nontransplanted trees (controls).
Discussion
The sizes of trees transplanted in this series of experiments were near the maximum that might be commercially transplanted by the mechanical tree spade of the size used. As a consequence, the trees often became severely water stressed, particularly during the summer. This is the type of controlled situation from which we could obtain information most applicable to that of transplanted urban trees (trees which periodically become severely stressed) and still provide a favorable soil environment for root growth. Poor soil conditions are common in the urban situation, and root regeneration rates may be considerably reduced from those reported in this study. The amount of root growth reported in this study is considered close to maximum for the species tested and the transplanting procedures used.
Root regeneration
Among the transplanted trees, the vertical distribution of regenerated fine roots was similar to the fine-root distribution of the control trees, taking into account the competition effect from vegetation. Figure 1 shows that root densities of the Norway maple and green ash controls were greatest in the upper 10 cm of soil. When grass competition occurred, root densities in the upper 10 cm were lower, as was found with ginkgo controls.
As might be expected in the first year after transplanting, densities of regenerated roots decreased with increasing distance from the root ball. However, as the diameter of the terminal root growth increased, the volume of soil exploited by the root system increased by a far greater percentage than the percentage of diameter increase. The regenerated roots at the periphery of the root ball constitute a major portion of the new root system of the tree and should be given primary consideration when watering and carrying out other maintenance procedures.
Root regeneration seems to be dependent on both the tree species and the size of the severed roots from which to initiate new growth. The severed roots (of all sizes) of Norway maple produced significant root regeneration after 1 year. Severed green ash roots were often larger than 4 cm in diameter and did not regenerate new roots even though smaller roots on the same tree showed substantial root initiation after the same 1-year period. This finding seems to partially explain the poor root regeneration of ash when compared with that of controls. Severed ginkgo roots were much less dense at the edge of the root ball due to the smaller trunk diameter of the trees. Also, the severed roots often died back up to 10 cm, and new roots initiated from the living cambial area. Consequently, when measured at the periphery of the soil ball, ginkgo root regeneration was low even though extensive fine-root initiation often developed.
Our studies seriously question the advantage of root pruning nursery trees to stimulate or cause to develop a more branched or compact root system confined primarily to the root ball area. When roots are pruned, regeneration occurs from the severed end. If roots are severed near this same location during transplanting, the entire new portion of the root system will be cut off and left in the nursery soil. It is probable that nothing is gained in tree vigor or future survivability by root pruning and in fact, there may be a negative effect, because root-pruning induces water and nutrient stress for one or two seasons prior to transplanting. The combination of root pruning in the nursery and later root pruning at the time of transplanting can induce susceptibility to fungus canker diseases and borer attack.
Carbohydrate assays
Results of these assays are in agreement with other reports of seasonal cycles in carbohydrate levels in wood tissue (6, 12). Our analyses indicate that roots of transplanted trees, a year after transplanting, have higher carbohydrate levels than nontransplanted trees have. This fact implies that photosynthates are being produced faster than they are being used. At first this may seem unlikely, but on further examination it becomes quite logical. Although the crown of the tree is somewhat reduced by pruning and the leaves are smaller, the root system has been more drastically reduced. Even if root regeneration occurs at a high rate, this probably does not require as much energy as is required for the growth of the original root system. Consequently, assimilates are likely to occur in excess. It is quite possible that these assimilates accumulate at the severed root ends and stimulate regeneration, just as has been reported in stem cuttings (13).
Previous workers have shown that root regeneration depends on carbohydrate levels in the tissues (7, 12), which depends on the physiological growth stage of the tree. At bud break, the elongating shoots act as a metabolic sink, resulting in a reduced flow of photosynthate to the roots. Consequently, root growth is slowed. Therefore, a poor time to transplant trees is just prior to bud break (3, 4). In this study, the Norway maples which were transplanted at the time of bud swelling had the poorest root regeneration of all maples transplanted in the four periods used in this study. Green ash transplanted simultaneously, had not yet broken dormancy and showed no reduction of root growth.
In late summer, 1980, carbohydrate levels in transplanted trees dropped, while rising in control trees. The summer was very dry, and it became difficult to maintain the proper soil moisture during the late summer months. The control trees, having normal root systems, were located in a dense stand where the soil was constantly shaded. This was not the case in the transplanted tree plot because of greater distances between trees and the consequent greater exposure to the drying effects of sun and wind. It appears that drought stress may be responsible for the late-summer decrease in carbohydrates. Parker (9) had shown this same effect of drought stress in controlled experiments on sugar maple seedlings.
Carbohydrate availability apparently is not the limiting factor for growth in the second growing season after transplanting. It is most likely that drought stress and/or mineral nutrient availability limit growth due to the reduced absorptive area of the root system.
Conclusions
Proper planting procedures and good maintenance of trees following transplanting are essential for good root regeneration. Many of the trees transplanted exceeded the recommended tree size limits for the equipment used, and moisture conditions were often less than ideal, yet the survival rate after 1 year was 97 percent. We attribute the transplanting success to minimum disturbance of the root ball and careful maintenance after planting. This success is in contrast to municipal street-tree plantings. Such plantings often consist of improperly handled nursery stock and are sometimes improperly planted and lack regular maintenance after planting. The losses in street, home, and highway tree plantings are often very high in the first year after transplanting.
Transplanted trees are subject to stresses resulting from the tremendous loss of root system. The successful reestablishment of these trees depends primarily on rapid root regeneration to maintain existing top growth. Root regeneration originates primarily at the severed ends of the roots cut during transplanting. For successful transplanting, the soil conditions must be favorable for root growth near the perimeter of the root ball. Regenerated fine-root systems are shallow in nature, with the highest root densities occuring within 10 cm of the soil surface. Competition with roots of grasses caused an inhibition of root growth in the upper 10 cm of soil.
The carbohydrate status of root tissue appears to be a limiting factor for root growth when trees are planted during the spring shoot-growth period. Root regeneration of Norway maples was decreased when they were transplanted during this period. Total nonstructural root carbohydrates were also lowest during this spring growth period. This was found to be the only period in which transplanting appeared to have a major impact on root regeneration.
Footnotes
↵1 Currently, Specialist, Michigan State University, East Lansing, Michigan
- © 1982, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.