
Abstract
The root system of a Norway maple affected by girdling roots was excavated, dissected, and examined microscopically. Cross-sectional area of vessels in the stem xylem affected by the girdle was only 10% that of unaffected wood. Rays in the stem wood were skewed and contained few pits. Bark on the girdled stems was compressed from a normal thickness of about 2.5 cm to 1 mm. The offending roots sustained slight compression of cells where they were in contact with the stem and appeared to remain functional. Thus, girdling roots apparently cause tree decline by reducing stem conductivity and radial communication between tissues.
Widespread planting of Norway maples (Acer platanoides) in the northeastern U.S.A. began in the 1930’s and increased during the 1950’s as first generation shade trees began to disappear and suburban communities grew. The species became popular because it was easy to propagate, grew rapidly, and was winter hardy on most sites. Norway maple was the major (and sometimes only) component of street tree plantings in many metropolitan and suburban sites during that era.
In the last 10 years, arborists and others who grow and maintain shade trees have voiced concern about a decline of Norway maple associated with self-girdling roots. Symptoms are reminiscent of the more widespread deterioration of sugar maple known as “maple decline,” i.e., leaf scorch, early fall coloration, reduced growth of leaves, twigs, and stems, and branch dieback (5,8). The shape of a Norway maple stem may also be indicative of the problem. Stems which, at the ground line, are cylindrical or which taper inward usually have girdling roots. Unaffected trees show normal root flares at the butt (4,6).
Arborists have attempted to restore health and vigor of trees suffering from girdling root injury by removing the offending roots, then fertilizing and watering (7). Limited success of these treatments raised questions about the nature of girdling root injury and the possibilities for remedial action. Our investigation was undertaken to clarify the nature of the injury.
Materials and Methods
A 25-year-old Norway maple that showed decline symptoms and was girdled by its own roots was selected for study. The tree was growing in sandy soil, the surrounding ground cover was turf, and the nearest obstruction to root growth was pavement 6 m from the stem. The stump was excavated, roots leading away from the butt were severed, and the remaining convoluted mass of basal stem and roots was collected. Cursory observations during excavation were noted, and numerous photographs of the then exposed roots were taken (Fig. 1).

Norway maple stump and root system with several girdling roots visible.
At the junction of the stem and the topmost girdling root, sections of the stem and root were excised and trimmed to blocks approximately 1 cm3. Blocks of ungirdled stem and root wood adjacent to the affected portions were also collected for comparison. Blocks were smoothed with a razor blade, air-dried for one week, coated with a thin film of gold, and viewed with a scanning electron microscope.
Presence of rays in some blocks was confirmed by observing thin sections stained with safranin (3).
Results
Upon excavation, it was evident that the topmost girdling root represented only part of a much more extensive problem. Four different major roots girdled various portions of the main axis from the ground line to a depth of about 20 cm. The extent of girdling by any single root ranged from 10-30% of the stem circumference. Each of the roots penetrated into the soil beyond the zone of girdling and appeared, based on color and texture of the tissue, to be wholly functional. None of the roots was grafted to the stem, and grafts between two roots were rare.
In order to interpret the effect of girdling on wood anatomy, it is necessary to appreciate the organization of cells in normal stem wood. Wood in Norway maple stems is diffuse porous. Large xylem vessels, the principal routes for ascent of water and nutrients, are distributed more or less uniformly throughout each growth ring (3) (Fig. 2A & B). Other vertically oriented cells are fiber tracheids. Their primary function is to give strength to the stem, but they are also sites for storage of food materials. Rays are oriented horizontally and extend from the bark in toward the center of the stem. Rays differ from other elements of the wood in that they are largely composed of living cells. They are avenues for the horizontal transport of photosynthate to and from storage in the fiber tracheids, and they supply nutrition to the vascular cambium to permit cell division and diameter growth (9).
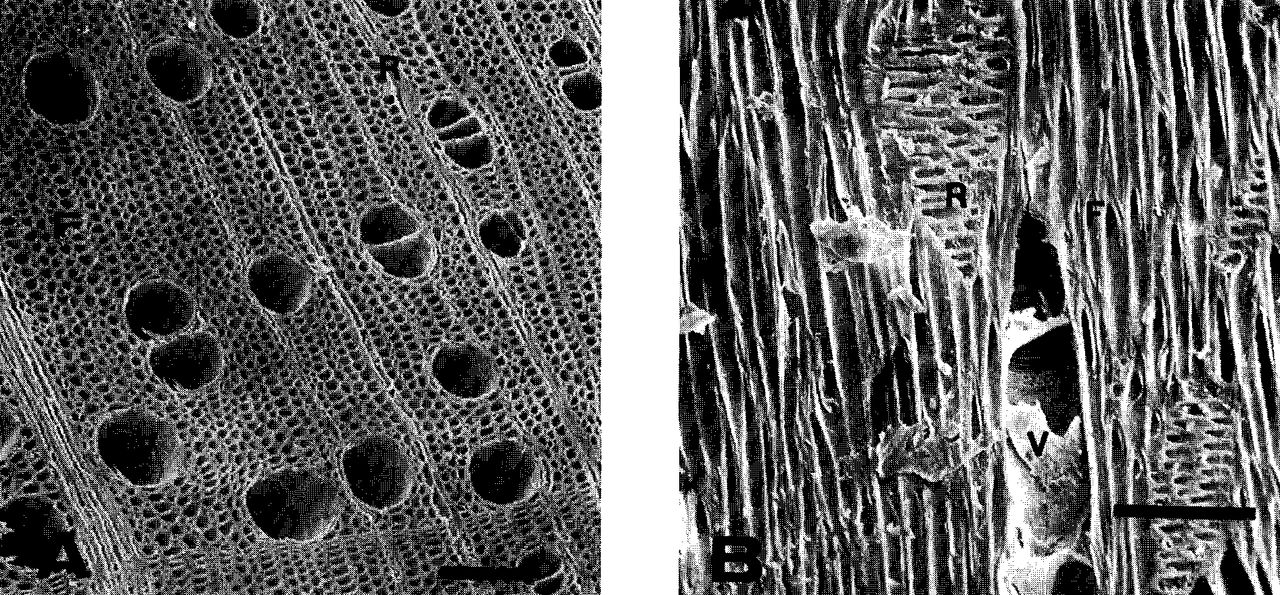
Transverse (A) and radial (B) views of normal Norway maple stem wood. V = vessel element. R = ray F = fiber tracheid. Scale of bar = 100jum.
Solutions move from one cell to another through simple pits which are little more than holes in cell walls (Fig. 3). Pits are numerous in walls of vessel elements and ray parenchyma where the two contact each other. They occur less frequently in the interior cells of rays and in the fiber tracheids (1).

Radial view of normal Norway maple stem wood. V = vessel element. R = ray. F = fiber tracheid. P = pits. Scale of bar = 100jum.
Measurements from four transverse sections taken from ungirdled portions of the stem indicated an average frequency of 41 (range: 26-54) vessels/mm2. Total cross-sectional area of vessels averaged 1885 (range: 1256-2205) jum2/mm2. Rays in our blocks appeared to contain only parenchyma and were up to 10 cells wide and 80 cells deep.
In that portion of the stem being girdled by a root, the structure of the wood and the orientation of cells were altered drastially (Fig. 4A & B). The mean number of vessels in three transverse sections was 31 (range: 26-36) vessels/mm2 and the average cross-sectional area of those was 104 (range: 12-268) μm2/mm2. Normal horiozontal rays were absent, but the vessels and fiber tracheids were interspersed with elliptical aggregations of cells which, when viewed in stained radial sections proved to be rays. The skewed rays had few pits, and those appeared to be occluded with cell wall depositions of unknown composition (Fig. 5),This pattern of abnormal anatomy occurred in tour annual rings immediately adjacent to the vascular cambium. Features of older xylem were normal.

Transverse (A) and radial (B) views of malformed Norway maple stem wood. V = vessel element. R = ray. F = fiber tracheid. Scale of bar = 100 jum.
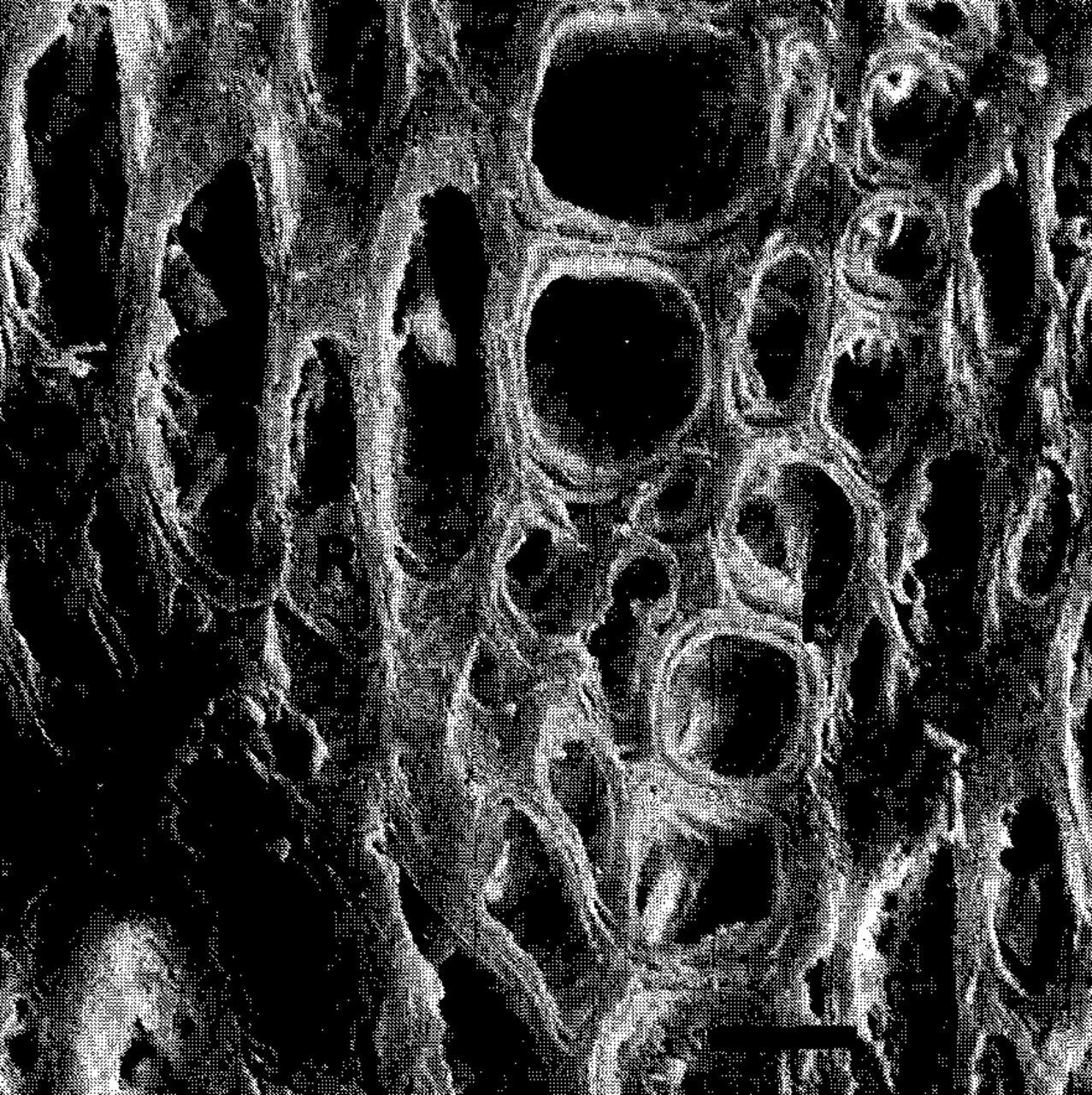
Transverse view of malformed wood to show debris and occlusions in the rays. Scale of bar = 5jum.
Root tissue, though slightly compressed near the point of stem contact, did not appear to be otherwise affected. Bark which varied from 1.5-3.0 cm thick in unaffected stems was compressed to a flaky aggregation of cells approximately 1 mm thick at the point of maximum contact between root and stem.
Discussion
The amount of self-girdling exhibited by the tree we studied was considerably more than expected at the outset. Superficial examination indicated that one girdling root was present, but after excavation we found four major roots encircling approximately 70% of the stem. Thus, to determine the potential for self-girdling, one must expose at least enough of a root system to account for all roots that would be buttress roots under normal circumstances.
The frequency of grafts between crossing roots or between roots and the stem was very low. This may simply have been an unusual characteristic of the particular tree we examined, but Graham and Bormann (2) were unable to find literature documenting self- or intraspecific grafting in Norway maple in their extensive review. If selfgrafting truly does not occur in this species, it would be especially sensitive to any contact with its own roots.
Xylem formed while under pressure from an adjacent expanding root has unusual anatomical characteristics which, to our knowledge, have not been documented previously. Not only is there a tenfold reduction in vascular conducting area, but rays are skewed such that they only faintly resemble those in normal wood, and their ability to function in a normal capacity is questionable.
Histological examination of the phloem was beyond the scope of this study, but cursory observations suggested that the phloem suffered as much if not more than the xylem. Thickness of girdled bark was up to 30 times thinner than the unaffected bark.
We suspect that girdling roots cause Norway maple decine by reducing (through pressure on the cambium) the size of the vessel elements and thus, the capacity of the vascular system to transport water and nutrients from roots to shoots. Flow of photosynthate to roots below the girdle is also apparently inhibited to the extent that they eventually die.
The ability of a girdled stem to recover following removal of the root could not be evaluated from our observations. A study spanning several years would be required to test this possibility. If roots below a girdle are, in fact, dead by the time crown symptoms appear, then removal of the offending root would be a trivial exercise. We are currently trying to relate symptom expression to extent of stem injury as part of our continuing studies on this problem.
The girdling root was relatively unaffected by its contact with the stem and was functioning to transport materials to other parts of the stem. Thus, severance of the root eliminates one major part of the root system and may actually hasten tree decline.
Footnotes
↵1 This research was supported in part by a grant from the International Society of Arboriculture Memorial Research Trust and by USDA, SEA-CRGO Grant No. 7900096 and NSF Grant No. PCM-7903263 awarded to J.R. Aist and H.W. Israel.
Presented at the annual convention of the International Society of Arboriculture in Hartford, Connecticut in August 1980.
- © 1981, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.