
Abstract
This paper points out the ephemeral nature of mycorrhizal short roots, influenced in their development by radiation, soil drainage, and eradicants. Emphasis is placed on rhizospheric and extramatrical mycelia as agents enriching soils in enzymes, chelates, and available nutrients. Inoculations of soil, seed, or stock with mycorrhiza-forming fungi without ascertaining the supply of major and minor nutrients and the soil’s freedom from toxic substances will inflict financial losses and undermine the program. Inoculation of biocide-impaired nursery soils usually necessitates addition of suitable energy material and adjustment of soil fertility. Suggestions are made for clarification of conceptual discrepancies relevant to tree-fungus symbiosis.
The symbiosis of tree and fungi is one of the most important relationships in the realm of plant physiology, especially in tree nutrition. As has been repeatedly demonstrated, seed of nearly all tree species planted in grassland soils, including those of very high fertility, usually yield singlewhorl seedlings which in one or two years die exhibiting signs of starvation. However, addition to the grassland soil of a minute amount of forest soil containing fungal symbionts of trees induces a most spectacular change: the starving, nearly dead seedlings recover from their lethargy and attain normal, often vigorous growth (Fig. 1).

Effect of inoculation of a prairie soil with mycorrhiza-forming fungi on the growth of 6-month-old seedlings of Monterey pine, Pinus radiata. A-Carrington (Plano) prairie silt loam; B-The same soil with addition of 0.2% by volume of humus-enriched horizon of Plainfield sand, a forest soil supporting jack pine and harboring mycorrhiza-forming fungi Cenococcum graniforme and Boletus luteus; C-Plainfield sand (After Wilde, 1968).
The symbiosis of trees and fungi has been investigated for over one hundred years. Yet, these studies have left many misinterpretations. This is particularly true of investigations conducted in artificial environments with little attention to soils and vegetative cover, especially its density. A very unfortunate disadvantage for many researchers of mycorrhizae was and still is their remoteness from soils of indigenous grasslands, the medium which infallibly reveals the presence or the absence of symbionts essential for the vast majority of trees and other lignophytes. Many years’ studies of the silvicultural importance of mycorrhizae, with Wisconsin prairie soils as a test medium, permit us to suggest essential corrections and simplifications of some commonly held concepts. All of these were established on the basis of repeated experiments and all of them can be verified by simple trials. We stress this point because some of our findings are in discord with credos expressed in recent literature.
Distribution of mycorrhiza-forming fungi
The fungal symbionts essential for the large majority of tree species, i.e., fungi of ectocellular and epirhizal development, are absent in prairie and other grassland soils which never supported trees (Wilde, 1958). Such soils are often located in the immediate neighborhood of forest stands and receive wind-blown spores of mycorrhizal fungi; yet they remain treeless because, in the absence of tree roots, mycorrhizal spores undergo the fate of unfertilized eggs. However, once established, mycorrhizal fungi preserve their symbiotic potential for a very long, indefinite period. Grasslands established on previously forested soils are not deficient in tree symbionts.
All forested or previously forested soils in all geographic regions, including soils subjected to severe fires, grazing, or cultivation, harbor fungi-forming mycorrhizae with all tree species. Our studies revealed them in Wisconsin bogmoss swamps, sand dunes, and soils which produced farm crops for a period as long as 60 years (Rosendahl and Wilde, 1942). According to the late Eric Bjorkman (1950), mycorrhizal fungi are present in Swedish soils which were under cultivation longer than a century. With the exception of soils subjected to prolonged inundation, such as those of abandoned beaver flowages (Wilde et al., 1950), there is probably not a single square foot of forested or previously forested soil in the world free from endocellular, ectocellular and epirhizal mycorrhiza-forming fungi essential for trees and other lignophytes. By and large, the statement, once forest soil, forever forest soil, i.e., soil harboring mycorrhiza-forming fungi (Wilde, 1954), preserves its validity. And outside of hydroponics and nutrient-enriched greenhouse sand cultures, there are no “nonmycorrhizal” or “autotrophic” trees.
The almost incredible perseverence of fungi essential for tree growth is best illustrated by the present distribution of vegetation in Wisconsin and Minnesota. In spite of clearcut logging, severe fires, and nutrient-depleted lands of abandoned farms, all previously forested soils that escaped man’s interference within the last 70 years now support naturally established forest stands. On the other hand, neighboring prairie soils remain treeless as they were during the many past centuries. The encroachment of forest upon prairie is largely limited to the narrow border strip penetrated by lateral roots of trees.
Inoculations with mycorrhiza-forming fungi
Introduction of mycorrhiza-forming fungi into grassland soils or soils scalped by grading does not present any particular difficulties. The first successful, large scale inoculation of chernozem soils, free from the essential tree symbionts, was accomplished about 80 years ago by Wissotsky (1902) who sprayed oak acorns with a thick suspension of forest topsoil. The same method is still the simplest and least expensive procedure for afforestation of grasslands by direct seeding. And it can be accomplished by the use of any forest soil. We achieved normal growth of many seed-planted coniferous and deciduous trees on prairie soils of Wisconsin by addition of a fraction of one percent of surface soil obtained from New Zealand, India, Italy, even Alaska permafrost and Wisconsin weathered limestone outcrops, or by spraying the seed with suspensions of these soils (Wilde, 1968).
Inoculation of grassland soils for production of tree nursery stock can be achieved by seed sprays, application of leafmold and humus-enriched surface soil from healthy forest stands, or by using the area during one or two years for transplanting seedlings obtained from a nursery producing healthy stock of a similar either ectocellular or endocellular mycotrophic make-up (Iyer, 1978b). Inoculation of hydroponic trees intended for planting on grasslands is accomplished by hilling them in any forest topsoil for a period of 2 or 3 weeks (Fig. 2). Planting of nursery-produced trees on grassland soils requires no inoculation with mycorrhizal fungi because there is no nursery stock of plantable size free from mycorrhizae of either ectocellular, endocellular, or epirhizal morphology.

Nine-month-old seedlings of white pine, Pinus strobus, raised for 2 months in a sterile nutrient solution and then transplanted to Carrington (Plano) prairie silt loam: A-seedlings transplanted directed from hydroponics to prairie soil; B-seedlings inoculated with mycorrhizal fungi by hilling them for 2 weeks in Plainfield forest sand prior to their transplanting to prairie soil.
In recent years, successful inoculations in the U.S.A. have been achieved with pure cultures of mycorrhiza-forming fungi, such as Pisolithus tinc-torius and Thelephora terrestris (Marx et al., 1978). However, the success of pure culture inoculations is dependent upon climatic conditions and compatibility of the exotic symbiont with native microorganisms, including mycorrhiza formers. In some instances, according to Marx et al. (lit. cit.), the pure culture inoculations are more successful following destruction of native microflora by fumigation. Moreover, successful introduction of a particularly effective symbiont, or any mycorrhiza-former into biocide-impaired nursery soil often necessitates addition of suitable energy material and adjustment of soil fertility required not only by the trees but also by the symbiont being introduced. In some cases, restoration of mycotrophy in eradicant-inhibited nursery soils was accomplished by application of fibrous papermill sludge and a nutrient-enriched suspension of leafmold (Iyer and Oilschlager, 1977).
An important detail pertinent to pure culture inoculations is that not all fungal symbionts of trees are wholly beneficial organisms. Thelephora terrestris, a fungus used recently for inoculation of nursery soils, forms large superficial blankets in forests eliminating all other plants by either toxic or antibiotic excretions. Thelephora laciniata, a close relative of Th. terrestris, is well known to European foresters as the most noxious of “smothering” fungi damaging and even killing nursery stock by enveloping stems of seedlings (Kavina, 1923).
Of equal importance is that mycorrhiza-forming fungi are not nitrogen-fixing organisms and cannot augment soil fertility. Hence, the introduction of a symbiont, regardless of its inherent effectiveness, can only promote utilization of the soil productive potential, but not increase it. No inoculation will promote the growth of trees on purely quartzitic sand, acid muskegs, soils lacking any essential major or minor nutrient, or soil containing toxic substances. The latter are of particular importance in industrial waste beds. Without a preceding thorough soil analysis, promiscuous inoculation with tree symbionts may result in waste of funds and undermine the entire program.
Specificity of fungal symbionts
Under natural conditions, existence of trees and other lignophytes is similar to that of lichens; it depends upon their inseparable union with symbiotic fungi. This union is foreign to segregation; a tree of any origin finds an acceptable fungal associate in any forested or previously forested soil not subjected to a prolonged inundation. And a distance of several thousand miles between birthplaces of the two symbionts presents no obstacle to their union. As was stated, we achieved normal growth of many seed-originated coniferous and deciduous trees on prairie soils by addition of a minute amount of surface soils obtained from distant parts of the world and thus demonstrated that specificity of mycorrhizal symbionts is a chimera. New Zealand provides the best illustration of the validity of this thesis. Both of its islands are dotted with exotic trees from Europe grown by the settlers from seed on soils which previously supported Nothofagus and other trees unknown in the rest of the world. The introduction of mycorrhizal symbionts was essential in New Zealand only for soils of grasslands.
Mycorrhizal short roots
In one form of tree-fungus symbiosis, fungal hyphae penetrate cortical tissues and produce, on the surface of roots, wartlike mycelial offshoots, named mycorrhizae or mycelial short roots. These mycelial appendages present an obvious display of the existing root-fungus union and they received a great deal of attention by the investigators of mycorrhizae. This concentrated effort may well be questioned because of the ephemeral, merely incidental nature of short roots.
Short roots are largely accessories of trees which receive a large supply of radiant energy, and their abundance is confined to trees of nurseries, greenhouses, and open stands. Shading by a dense canopy drastically reduces and even completely suppresses the development of mycorrhizal short roots (Fassi, 1967).
To demonstrate to our students the imaginary importance of the short roots in the growth of natural stands, we conducted at different times during the growing season, repeated, careful excavations of 3 to 10-year-old seedlings of red pine, Pinus resinosa,and white pine, P. strobus. Most of these excavations were performed in the well-stocked, virgin stand of red and white pines, located in the University of Wisconsin Finnerud study area in Oneida County. With practically no exceptions, excavated trees revealed no mycorrhizal short roots, but only superficial mycelia and mycelia-agglutinated terminal clusters (Fig. 3). Identical root morphology was disclosed in naturally reproduced seedlings of dense stands of ponderosa pine, Pinus ponderosa, and Douglas fir, Pseudotsuga menziesii, of Idaho and Oregon (Benson and Iyer, 1978).

Roots of naturally reproduced seedlings of red pine, Pinus resinosa, and white pine, Pinus strobus, exhibiting grumose, mycelia-agglutinated clusters, but no mycorrhizal short roots. Randomly excavated trees from a soil supporting a dense, virgin stand of red and white pines in the Finnerud study area of the University of Wisconsin in Oneida County, Wl.
The influence of radiation on the development of short roots is also revealed conspicuously by their relative abundance in nursery stock of different ages and corresponding density. The maximum occurrence of short roots is usually in 1-year-old seedlings exposed to radiation, and the minimum in fully-stocked 3-year-old seedlings, the foliage of which receives only a small fraction of full sunlight (Iyer, 1978a). Particularly great differences in the abundance of short roots are also observed in nursery stock located on the border and in the center of nursery beds (Fig. 4).

Effect of radiation on the development of mycorrhizal short roots of 2-year-old seedlings of red and white pines. A and C: mycorrhiza-endowed roots of trees lifted from the borders of nursery beds; B and D: mycorrhizadeficient roots of trees lifted from the center of nursery beds. Wilson state nursery of Wisconsin (After Iyer, 1978).
Disappearance or radical reduction of mycorrhizal short roots is also prevalent in dense forest plantations. In the course of the Wisconsin statewide survey of coniferous plantations (Wilde et al., 1965), jet excavations of many red and white pine sample trees revealed no, or near absence of short roots (Wilde and Iyer, 1962; Wilde, 1967). All our observations left the impression that mycorrhizal short roots are symptoms of a measles-like disease, inflicted by exposure to full sunlight.
Epirhizal mycelia and terminal mycelial clusters
While the past investigations of mycorrhizae devoted the lion’s share of attention to mycorrhizal short roots, they have largely neglected the composition and effects of mycelia which cover the roots with mantles, mycochlenes (Peyronel, 1922), and pellicles, mycochlamides, and mycelia which form agglutinated, terminal clusters, mycoplasts (Wilde and Lafond, 1967). The symbiotic or mycotrophic nature of superficial and terminal mycelia cannot be doubted. Their very growth on the surface and at the tips of roots testifies that they are beneficiaries of trees, receiving from them absolutely essential and otherwise unprocurable carbohydrates. In turn, these mycelia are benefactors of their host plants for they enrich the soil in enzymatic and chelating compounds effecting the solubilization of available nutrients (Leaf, 1957; Spyridakis et al., 1967;Voigt, 1969; Wilde et al., 1978). The sym-biotrophic effect of epirhizal and terminal mycelia is convincingly revealed by trees of closed canopy deprived of direct sunlight and producing no mycorrhizal short roots. In this connection, it should not be forgotten that some, perhaps a great many, “contact” fungi such as Trichoderma viride produce, by their mere presence in the proximity of tree roots, the same growth effects as do root-penetrating fungi (Wilde et al., 1956). Trichoderma also provides a very instructive illustration of the antiseptic role of epirhizal mycelia; its excretions of antibiotic substances eliminate nearly all other soil microorganisms (Yatazawa et al., 1960).
Among the variants of epigenous mycelia, terminal clusters, or mycoplasts, provide the best opportunity for detailed investigations. This form of rhizospheric union of fungi and tree roots permits its easy isolation, microscopic examination, and analysis. The previous neglect of these essential components of tree roots has been largely due to their fragile nature and very weak connection with the roots. In lifting trees, the clusters are usually broken from the fine rootlets and left in the ground, or detached from the roots by shaking or washing in an effort to remove adhering soil particles.
Depending on the nature of the soil, the round or oval mycelia-agglutinated clusters vary in size from a few mm to about 3 cm (Fig. 5). They consist of an intimate aggregation of soil particles and fine rootlets enmeshed in a network of fungal mycelia. Mycroscopic examination often reveals fragments of silicate minerals in a state of advanced weathering. The clusters formed within the soil layers enriched in humus include finely dispersed organic matter (Fig. 6).

Mycelia-agglutinated rhizospheric root clusters or mycoplasts of a naturally reproduced 5-year-old white pine seedling.

Cross-section of a mycelial root cluster comprising an aggregate of rootlets, fungal mycelia, soil particles, and dispersed organic matter (10 ×).
Analysis of roots and their terminal clusters of 3-to 7-year-old seedlings of white and red pines included determinations of catalytic potential, reaction, exchange capacity, and exchangeable bases (Wilde et al., 1979). These analyses showed that the enzymatic content of the active fraction of clusters is about 20 times as great as that of their parent rootlets. The clusters also revealed about four times as great a cation exchange capacity and a much higher content of exchangeable cations in comparison with surrounding soils (Wilde et al., 1978).
These results suggest the following concludions. The high concentration of enzymatic substances in the terminal clusters testifies to a high biological activity. The growth of microorganisms, predominantly mycelia of non-parasitic fungi, in these root appendages is dependent on the availability of otherwise unprocurable carbohydrates from the host plant. The observed production of organic colloids and the subsequent concentration in the vicinity of the roots of exchangeable cations indicates a gradual release of these nutrients in accordance with requirements of the host plant. Under conditions of dense forest cover, mycelial clusters most likely perform the function that mycorrhizal short roots perform under high light intensity. It is probable that mycelia of the clusters exert an aseptic influence by release of antibiotic substances.
The other important factor in the production of enzymes, chelates, and available nutrients is the activity of extramatrical mycelia. Their enormous network extends through the entire area of root distribution and, at times, produces so-called “fairy rings” of mushrooms. Recent investigations have disclosed that external mycelia of grafted roots of stumps continue their delivery of nutrients to trees spared in partial cuttings (Dosen and Iyer, 1979).
A tree lifted from the ground is usually only a part of an organism because an essential component, the nourishing mycelium is left in the soil.
Mycorrhizal short roots and the quality of nursery stock
The notion that mycotropic capacity and the value of nursery stock are expressed by the relative abundance of mycorrhizal short roots is erroneous. Only in some instances does the quantity of short roots serve as an indicator of the nursery stock quality; in other instances, the abundance of short roots is of questionable or no significance. Nursery stock with superabundant short roots may have a very low performance potential; vice-versa, stock with sparse short roots may be excellent planting material.
During recent years, suggestions have been made that high fertility of nursery soils depresses the development of mycorrhizal short roots and lowers the survival of outplanted trees. In consequence, attempts have been made to record the root morphology of nursery stock by counting short roots and expressing their abundance as percentages of feeder roots.
As was reported by Benson and Iyer (1978), under conditions of an accepted density, 2-year-old nursery seedlings of ponderosa and lodgepole pine, Douglas fir, and Engelmann spruce exhibited adequate development of short roots in spite of an exceptionally high level of soil fertility including 400 lbs/a of available phosphorus pentoxide and 500 lbs/a of available potassium. By and large, a reduction in the abundance of short roots is not effected by high soil fertility, but by the density of nursery beds, correspondingly diminished radiation of the foliage, and by phytotoxicity of potent eradicants.
Counting short roots, especially if preceded by staining, is a tedious and costly procedure and the percentages are not acceptable values in this type of analysis; they are likely to provide a totally misconstrued picture of root morphology and misleading information on the performance potential of planting stock.
In appraisals of nursery stock quality, the relative abundance of short roots may have significance only as a supplement to more important attributes of trees, such as color of the foliage, root-top and height-diameter ratios, specific gravity of stems, and foliar composition. In rapid surveys, the development of short roots can be recorded in accordance with the following scale: superabundant-3, abundant-2, sparse-1, absent-0. In more exacting appraisals, the relative abundance of short roots along with other forms of mycorrhizal fungi can be rapidly estimated on the basis of the enzymatic content of rootlets smaller than 2 mm in diameter by manometric analyses (Wilde et al., 1979). These analyses reflect the relative abundance of short roots and also provide information on the biological activity of the nutritionally most effective part of the root systems of trees.
According to the results obtained in a survey of stock produced in five large nurseries of the Lake States region (Iyer, 1978a), the catalytic potentials in fully-stocked jack, red and white pines, and white spruce were expressed by the following averages: one-year-old seedlings, 55± 9.1 mm Hg/g; two-year-old seedlings, 42 ± 8.5 mm Hg/g; three-year-old seedlings, 26 ± 7.2 mm Hg/g. These averages indicate a linear relationship between catalytic potential and age with a standard error of estimate in the proximity of 8 mm Hg/g. A similar relationship was noted for incidence of short roots. Figure 7 illustrates the significance of the catalytic potential by featuring the extremes encountered in the development of roots of nursery stock. Experience in determination of catalytic potential may help nurserymen appraise the direct and mycotrophic capabities of roots by ocular estimates.

Catalytic potential of roots of 3-year-old white pine seedlings. A-seedlings with inadequately developed roots; catalytic potential of 12 mm Hg; B-seedlings with abundant development of feeder rootlets and mycorrhizal short root; Cp of 63 mm Hg.
Mycorrhiza and eradicants
The great concern about the effect of high fertility of nursery soils on the development of mycorrhizae is rather out of place at this time when nearly all nursery soils are treated with potent, often destructive biocides (Iyer, 1963; Iyer et al., 1969). Some of these chemicals at large concentration, or partial detoxification, eradicte both ectocellular and rhizospheric symbionts and depress the growth of trees (Fig. 8). Such effects may be particularly expected in soils with impeded drainage (Morby et al., 1978).
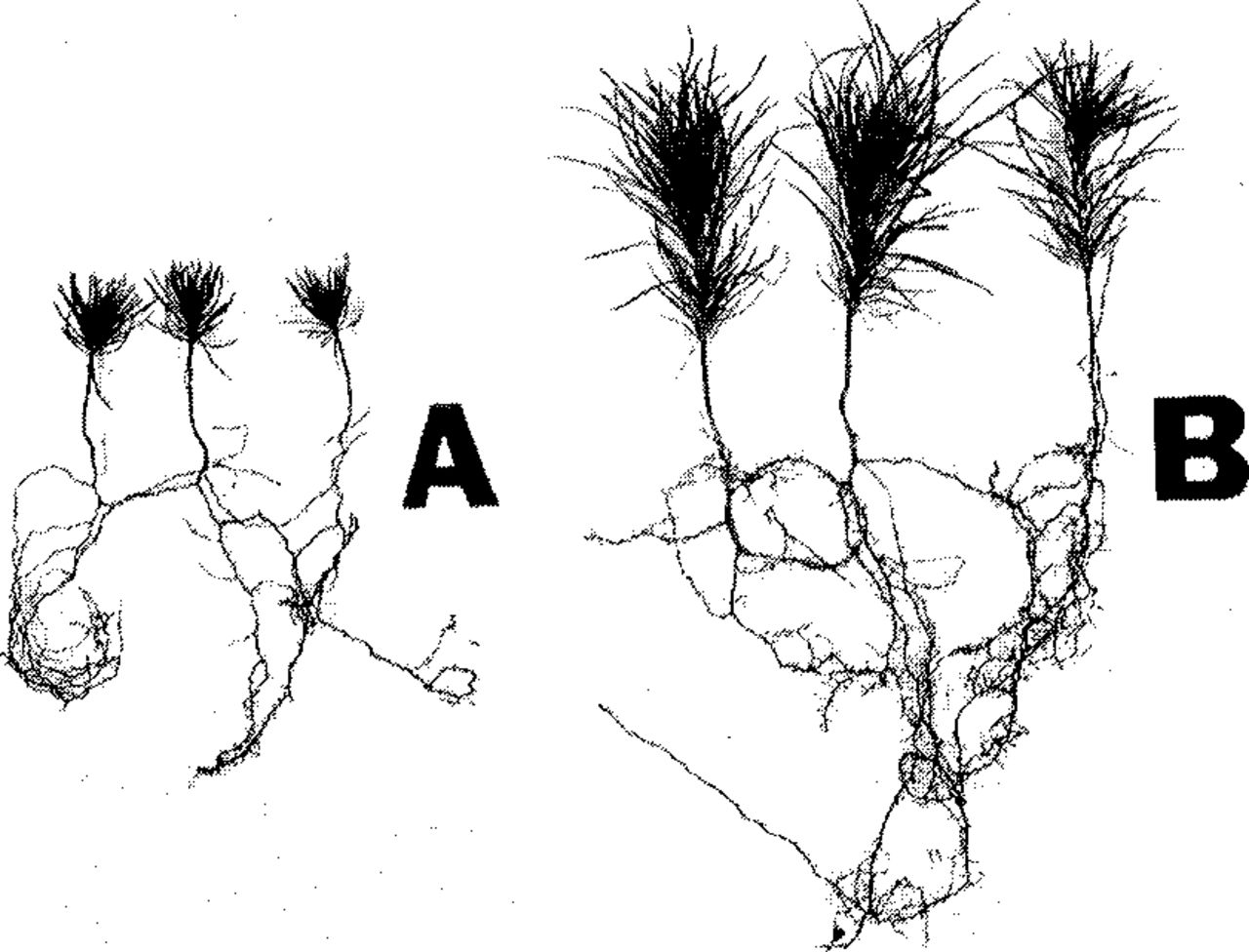
Red pine seedlings, in the second year of their growth, raised in depressions with highly concentrated Mylone (DMTT) herbicide (A); normally developed seedlings on elevated parts of the same nursery bed (B), exhibiting initial development of mycorrhizal short roots (Griffith state nursery of Wisconsin).
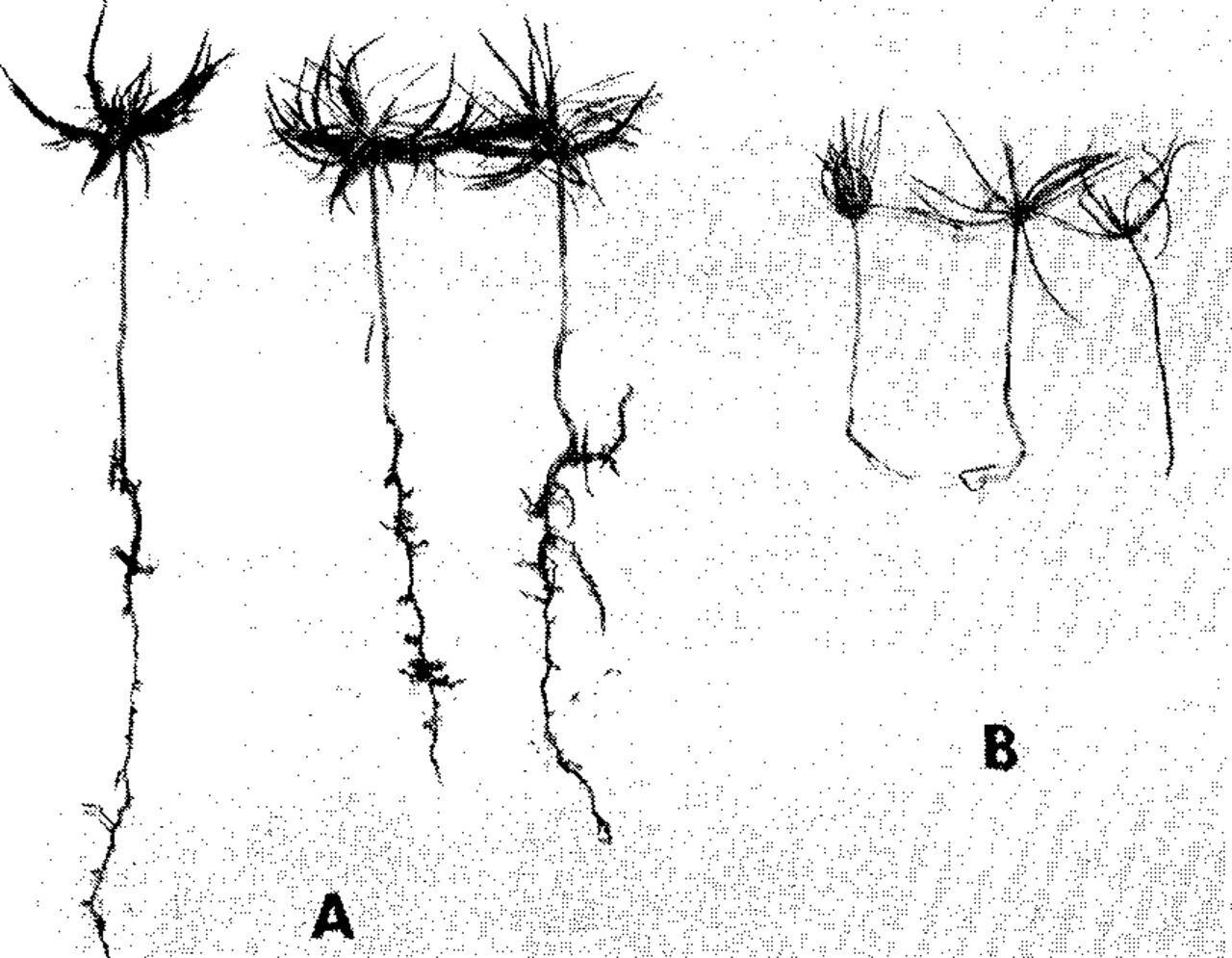
On the other hand, when eradicants are applied at reduced concentration, they may still annihilate mycorrhizal short roots and yet induce an exuberant, highly harmful growth stimulation of tree crowns and impart to nursery stock calamitous top-root ratios (Fig. 9) and succulent tissues (Iyer and Wilde, 1965).

Three-year-old red pine seedlings raised in eradicant-free soil (A) and in soil that received 60 lbs/a of Vapam (SMDC) fumigant (B). Note abnormally large crowns, but drastically reduced roots (Hayward state nursery of Wisconsin).
The destruction of mycorrhizae, or the entire root system of young seedlings, may at times be caused not only by toxic chemicals, but also by biotic eradicants, such as green manure of sorghum-Sudan hybrids, containing hydrocyanic acid (Fig. 10).
Effect of sorghum-Sudan green manure on the growth of 9-week-old white pine seedlings. A-normally developed seedlings raised in untreated sandy soil; B-seedlings raised in a similar soil treated with green manure of sorghum-Sudan hybrids and exhibiting “burning” of roots similar to that produced by lead arsenate.
Under present conditions, nursery stock production must be concerned with the development of all roots, be they short, long, or intermediate, and also with a number of other external and internal attributes of trees (Wilde et al., 1979).
Acknowledgment
Research supported by the College of Agricultural and Life Sciences, University of Wisconsin-Madison, and the Wisconsin Department of Natural Resources.
- © 1980, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- Distribution of mycorrhiza-forming fungi
- Inoculations with mycorrhiza-forming fungi
- Specificity of fungal symbionts
- Mycorrhizal short roots
- Epirhizal mycelia and terminal mycelial clusters
- Mycorrhizal short roots and the quality of nursery stock
- Mycorrhiza and eradicants
- Acknowledgment
- Literature Cited
- Figures & Data
- Info & Metrics
- References
Related Articles
Cited By...
- No citing articles found.