
Abstract
Background: Emerald ash borer (EAB), Agrilus planipennis, is an invasive wood-boring beetle that threatens ash trees (Fraxinus spp.) in the urban forest. Methods: We compared the efficacy at which 3 different injection systems deliver emamectin benzoate (EB) to the leaf canopy and protect trees. Two of the systems, Arborjet Tree I.V.TM and Rainbow Ecoscience Q-ConnectTM, used sixteen ports/m of trunk circumference, whereas the third system, Brandt enTREE® EB, used eight. Results: The two systems with more injection ports provided more uniform delivery of insecticide to the leaf canopy. Although all 3 injection systems provided excellent control for the first 2 years, only the 16-port injection systems provided protection for up to 3 years. The number of injection ports affected insecticide delivery because the sectorial structure of the ash vascular system limited lateral product diffusion. Additionally, over the course of an 8-year study, we found trunk injections of EB made in the spring provided better control of EAB than fall injections. Protection afforded by a 2013 application in our study failed by 2016 as EAB populations reached their peak. A second application in 2016 extended protection past the peak of EAB abundance through 2019. Conclusions: We conclude that when the same dose of EB is delivered, efficacy of the application is influenced by the number of injection ports used in the injection system, time of application, and the status of the local EAB population. Arborists need to be aware of these factors when planning their EAB management program.
INTRODUCTION
The emerald ash borer (EAB; Agrilus planipennis Fairmaire)(Coleoptera: Buprestidae), a key pest of urban ash trees, is arguably the most economically important insect pest to threaten North American forests (McCullough 2019; Ward et al. 2021). Since it was detected in 2002 near Detroit, MI, EAB has killed hundreds of millions of ash (Fraxinus spp.) trees across much of the eastern United States and parts of Canada and has spread to 35 states and 5 Canadian provinces (Emerald Ash Borer 2021). All North American ash species examined to date are susceptible to EAB attack; however, blue ash (F. quadrangulata Michx.) shows some resistance (Anulewicz et al. 2008; Tanis and McCullough 2015). The larval stages of EAB feed on phloem and cambium tissue and eventually scar functional xylem, cutting off the vascular system of their ring porous hosts (Herms and McCullough 2014). This injury kills limbs that then become dry and structurally unsound (Persad et al. 2013; Persad et al. 2019). Entire trees can be killed within 3 to 6 years after infestation, depending on local EAB density, tree size, and species (Cappaert et al. 2005; McCullough 2019).
Emerald ash borer adults emerge from ash trees in late spring, leaving behind characteristic D-shaped exit holes, and stay active throughout the summer. Adults feed on leaves for 1 to 2 weeks before mating and ovipositing in cracks and crevices of bark on the trunks and branches of ash trees (Wang et al. 2010). Although most eggs are laid within 100 m of the adult emergence site (Mercader et al. 2009), females prefer to oviposit on stressed and previously infested trees. This oviposition preference suggests their actual disperal is much shorter (McCullough et al. 2009a; McCullough et al. 2009b). After making its characteristic serpentine galleries, the last instar larva bores approximately 1 cm into sapwood of younger trees or the outer bark of mature trees and folds over itself to spend the winter as a prepupa (Poland and McCullough 2006). Prepupae develop into pupae during the spring and emerge several weeks later as adults. Larvae may require 1 season to develop into prepupae in trees that are unhealthy but can take more than 1 season to reach this stage in healthy, vigorous trees (Yu 1992; Tluczek et al. 2011).
The demise of ash trees is particularly important to urban forests where they were widely planted as street trees prior to the arrival of EAB because of their form, fast growth, tolerance to a wide variety of growing conditions, and apparent resistance to pests and diseases (APHIS 2003; Cappaert et al. 2005; Poland and McCullough 2006; Raupp et al. 2006). In addition to their aesthetic value, urban ash trees provide ecosystem services such as water filtration, improved air quality, carbon sequestration, and microclimate regulation (McPherson et al. 1997; Cregg and Dix 2001; Willis and Petrokofsky 2017). According to a 2010 model, the estimated cost of treating, removing, and replacing approximately 17 million urban ash trees across 25 northeastern states would exceed $10 billion by 2020 (Kovacs et al. 2010).
Applications of insecticide can protect urban ash trees from EAB (Herms et al. 2019), and, of the recommended products, emamectin benzoate (EB) is most acutely toxic to larvae (Poland et al. 2016). In fact, 1 year after trees were injected with this insecticide, over 75% of adults feeding on leaves of these treated trees died after 1 day of feeding and 100% were killed after 4 days (McCullough et al. 2011). This insecticide is most effective at protecting trees that have a healthy vascular system and are asymptomatic or in the early stages of EAB infestation (Flower et al. 2015; Bick et al. 2018; McCullough et al. 2019; Sadof et al. 2021). While the above studies suggest a single injection affords 3 years of protection, others report a duration of effectiveness between 2 to 4 years (Smitley et al. 2010).
A wide range of economic analyses suggests that protecting healthy ash trees from EAB with insecticides can be more cost effective in an urban forest than simply removing trees as they die and replanting them with resistant species. When these analyses assume that only individual trees treated with insecticide will be protected, economic advantage depends on treating trees once every 3 years (Kovacs et al. 2010; Vannatta et al. 2012; Kovacs et al. 2014; Sadof et al. 2017). In contrast, when these analyses include assessing areawide management strategies, treatments are required every 2 years to slow the mortality of nearby untreated trees (McCullough and Mercader 2012; Kibiş et al. 2020; Bushaj et al. 2021; Sadof et al. 2021). The success of all these programs hinges on deploying insecticide in a manner that reliably protects trees.
Tree injection has emerged as an efficient way to deliver plant protectants to trees (Berger and Laurent 2019). Numerous injection systems are available that vary in the number of injection points and whether or not ports are plugged to prevent leakage of product from injection sites. Arborists commonly inject EB into the base of a tree in spring to infuse the canopy with active ingredient before adult EAB emerge and begin to feed and mate, and females oviposit. In addition to killing adults feeding on leaves, this application kills first and second instar larvae that develop in early summer. Some arborists, however, inject EB in the fall to kill late-stage larvae that are actively feeding and also target larvae and adults the following spring. In this study, we evaluated the extent to which 3 commercially available injection systems influence the uptake and persistence of EB in the canopy of mature ash trees and compared their ability to protect trees from EAB after injection. In addition, we conducted a long-term study to examine the capacity of spring and fall injections to provide persistent protection along the invasion wave of EAB.
MATERIALS AND METHODS
Uptake Study
Uptake and persistence of EB were compared between trees treated using 3 different injection systems. Twenty white ash trees (Fraxinus americana L.) located at the campus of Culver Academy in Culver, IN, USA, were used in the study. Trees in this study were growing in open, park-like conditions within 200 m of unmanaged forest stands. These trees ranged from 40 to 64 cm DBH (trunk diameter 1.37 m above the soil) with a mean of 49.12 ± 8.79 cm. All 20 trees were previously treated with annual imidacloprid soil drenches beginning in the spring of 2010. It was not feasible to use trees without recent history of insecticide application due to the high EAB pressure in the area; however, these prior treatments were unlikely to significantly affect this study because of the relatively short window of protection provided by imidacloprid treatment (Bick et al. 2018; McCullough et al. 2019). Although we did not measure EAB abundance, we noted that the EAB population was high enough on the date of application for us to observe adults crawling on the tree trunks and spray equipment.
On 2016 June 16, 20 trees were grouped by location into 5 replicate blocks of 4 trees. Each tree within a block was randomly assigned to 1 of 4 treatments (Table 1). Treatments consisted of an untreated control and 4% active ingredient EB injected with either Q-ConnectTM (QC, Rainbow Treecare Scientific Advancements, Minnetonka, MN, USA), enTREE® EB (enTREE, Brandt Consolidated, Inc., Springfield, IL, USA), or Tree I.V.TM (Tree I.V., Arborjet, Inc., Woburn, MA, USA). ArborMectinTM (Rainbow Tree-care Scientific Advancements) was the EB product used in QC and enTREE injection systems. TREE-äge® (Arborjet, Inc.) was the EB product used in the Tree I.V. injection system.
Characteristics of injection systems and products used to inject undiluted 4% emamectin benzoate into white ash trees (F. americana) in Culver Academy and Eagle Creek Park, IN, USA.
All trees were injected according to manufacturer instructions with undiluted insectide on 2016 June 16 at a rate of 0.2 g active ingredient per 2.5 cm DBH and adequate soil moisture (Table 1). Injection holes were made with new helical drill bits and equally spaced around the trunk and placed in root flares when available. Other injection sites were placed within 10 cm of the soil line, avoiding basal defects or flattened sections of the trunk where translocation could be impeded by a girdling root. Injection lines were primed prior to injection to bring the liquid within an inch (2.5 cm) of each injection port. These ports were then monitored during injection to assure that they did not leak and the blue-colored insecticide flowed freely. Degree days accumulated from January 1 to the day of application (Table 2) were calculated from data obtained 20 km away in a Plymouth, IN, USA, weather station (PADE 2021) using the double sine method with a lower threshold of 10 °C (IPM 2021).
Conditions during injection treatments of 4% emamectin benzoate applied to F. americana in Culver Academy and Eagle Creek Park, IN, USA.
To track translocation of EB from injection sites into the canopy, we measured the concentration of EB present in leaves 1 week and 1 month following injection. Concentrations of EB in branch phloem and xylem were not measured. Using a weighted throw line, 5 leaf samples were collected from each tree on 2 sample dates 1 week and 1 month after treatment: 2016 June 23 and 2016 July 16. To ensure that these samples were representative of the entire canopy, 1 sample was collected from the top of the canopy, 2 samples were collected from opposite sides of the middle canopy, and 2 samples were collected from opposite sides of the lower canopy. Leaf samples were placed on ice immediately upon collection until returning to the lab where they were stored at −20 °C until EB could be extracted and then quantified at the Bindley Bioscience Center (Purdue University, West Lafayette, IN, USA). Leaves from 1 control tree were not collected, therefore the leaves of that tree were excluded from analysis of foliar EB.
Emamectin Benzoate Quantification
Sample preparation was conducted according to the methods of Burkhard et al. (2015). In brief, approximately 0.9 g of frozen leaf tissue from each of the 5 samples was placed into 7 mL Precellys lysing tubes along with 2.0 g of 2.8 mm zirconium beads (approximately 30 beads) and 4 mL acetonitrile. Leaves were homogenized in a Precellys tissue homogenizer for 3, 20-second pulses at 6,000 rpm alternated with 30-second pauses. After homogenization, samples were transferred to 15-mL conical falcon tubes and 6 mL of acetonitrile and water (95:5) were added to bring the final volume to 10 mL.
A 1.2-mL aliquot was removed and added to a QuEChERS extract tube containing 0.24 g MgSO4 and 0.06 g sodium acetate. Tubes were shaken and centrifuged at 2,500 × g for 2 min. The supernatant was transferred to a dispersive SPE tube (Agilent Part No. 5982-5022), shaken, and centrifuged at 2,500 × g for 2 min. The supernatant was transferred to a microcentrifuge tube and dried in a vacuum concentrator. Samples were reconstituted in 75 μL of acetonitrile and water (80:20), sonicated, and centrifuged at 16,000 × g for 8 min, and the supernatant was transferred to HPLC autosampler vials (HPLC-MS/MS Analysis). Emamectin benzoate levels were quantified by HPLC-MS/MS, based on the protocol in Durdent (2007). Separation was performed on an Agilent Rapid Resolution 1200 HPLC system using an Agilent Zorbax SB-Phenyl (2.1 × 50 mm, 3.5 μm) column. Mobile phase A was water and acetonitrile (90:10), and mobile phase B was water and acetonitrile (10:90), both with 0.1% formic acid and 50 mm ammonium acetate. Initial conditions were 50:50 A:B, followed by a linear gradient to 0:100 at 5 min and held until 7 min. Column re-equilibration was performed by returning to 50:50 A:B at 7.5 min and held until 10 min. The column flow rate was 0.4 mL/min, and the retention time for EB was 2.6 min. Analytes were quantified by MS/MS utilizing an Agilent 6460 triple quadrupole mass spectrometer with electrospray ionization. Quantitation was based on multiple reaction monitoring. Electrospray ionization positive mode was used with a transition of 886.5 to 158.0 (quantifier) and 886.5 to 302.0 (qualifier), with a collision energy of 32 V. A fragmentor energy of 100 V and a dwell time of 100 ms were used. Source parameters were as follows: nitrogen gas temperature = 325 °C and flow rate = 10 L/min, nebulizer pressure = 241 kPa, sheath gas temperature = 250 °C, sheath gas flow rate = 7 L/min, and capillary potential = 4.0 kV. All data were collected and analyzed with Agilent MassHunter B.03 software. Quantitation was based on a 5-point standard curve, with concentration range from 0.25 to 200 ng/mL, by spiking EB into untreated leaves. Standard curves were fit to a linear function with a 1/x curve fit weighting. Correlation coefficients were > 0.97.
Effects of Injection Method on Efficacy
To measure and compare the capacity of each injection system to protect ash trees from EAB, percent canopy thinning of each tree in our study was visually estimated to the nearest 10% (0% is full canopy, 100% is a dead tree with no leaves) on 2016 July 19, 2017 September 12, 2018 August 17, and 2019 August 22 (Rebek et al. 2008; Smitley et al. 2008). Ash canopy thinning has been correlated with EAB larval density, so visual canopy ratings provided us with a rapid, noninvasive, and accurate method to quantify the protection provided by each treatment over time (Smitley et al. 2008). Mean canopy thinning was compared each year in a randomized complete block design.
Effects of Application Timing on Duration of Efficacy
The efficacy of spring (mid- to late June) and fall (late September to early October) applications of EB to protect against EAB were evaluated on newly symptomatic ash trees in Eagle Creek Park in Indianapolis, IN, USA. Trees used in this study were mostly located within the forest and all within 100 m of the forest edge. The 30 F. americana trees with an average trunk diameter of 101.4 ± 4.79 cm (39.22 ± 1.86 in) were grouped into 3 sets of 10 replicate blocks. Within each block, trees were randomly assigned to receive injections of TREE-äge® insecticide using the previously described plug system at 3-year intervals in the spring or fall, and untreated trees served as controls (Table 1). In addition to following previously described injection protocols, the second round of injections was placed at least 2 in (5 cm) away from previously placed injection ports. The thinning of ash canopy was evaluated yearly as described previously on 2013 September 11, 2014 August 8, 2015 August 13, 2016 August 3, 2017 August 14, 2018 August 16, 2019 August 13, and 2020 August 31. The degree days (base 10 °C) on the date of each application (Table 2) were accumulated as described previously from data collected at the Eagle Creek Airport 5 km away from the study site.
Statistical Analysis
We estimated the evenness of insecticide distribution from the proportion of leaf samples from each tree that contained more insecticide than the untreated controls. The average concentration of EB across the 5 leaf samples collected from each tree was used as the single insecticide concentration value of each tree in our analysis. To meet the assumptions of normality and heteroskedasticity, concentration data were log10 transformed and proportions of infested leaves were arcsine square root transformed for all analyses. Effects of each injection method on the uptake of insecticide and its persistence were analyzed in a repeated measures analysis of variance using PROC GLIMMIX (SAS 9.4 Institute 2020) with a controlled spherical covariance structure. Model fit was checked with a restrictive maximum likelihood method using the Akaike information criteria. Differences in mean concentrations and proportions of leaves with detectable EB were compared in post hoc least squares means procedure using Tukey’s Honest Significant Difference test to control for experiment-wise error.
Efficacies of each treatment or injection method on the protection of trees at the Culver Academy and Eagle Creek Park sites were also evaluated by using a repeated measures analysis of variance using PROC GLIMMIX (SAS 9.4 Institute 2020) with a controlled spherical covariance structure. Model fit was checked as previously described. Mean canopy thinning per treatment was compared during each evaluation date in post hoc least squares means procedure using Tukey’s Honest Significant Difference test to control for experiment-wise error.
RESULTS
Uptake Study
The proportion of leaf samples with EB residues greater than the control varied significantly across treatments (F(2,12) = 4.04, P = 0.045) but not sample dates (F(1,12) = 1.06, P = 0.324)(Table 3). Trees treated with the enTREE system had 64% of the leaf samples with more EB than the control 1 month after injection. This percentage was significantly lower (P = 0.0314) than the percentages of emamectin-infused leaf samples collected from trees treated with the QC (96%) and Tree I.V. (96%) systems. Concentrations of emamectin residues varied significantly across treatments (F(3,16) = 19.08, P < 0.0001)(Table 3). Insecticide residue in the untreated trees was not significantly different from 0 ppb (P = 0.96), while leaves of trees treated with insecticide had more EB than the control (P = 0.0021). There was also significantly more emamectin in leaves 1 month than 1 week after treatment (F(1,16) = 18.31, P = 0.0006), a trend that did not change across treatments (F(3,16) = 0.38, P = 0.76). Experiment-wise comparisons between insecticide residues in trees injected with the enTREE system did not differ between the QC (P = 0.216) or the Tree I.V. systems (P = 0.17). Nevertheless, the mean concentration of insecticide was 51% lower in leaves of trees treated with the enTREE system than the other 2 injection systems. The difference between mean concentrations of EB residues in leaves from trees treated with Tree I.V. and QC systems was only 5.1% and not significant (P = 0.99).
Means and standard errors of emamectin benzoate (EB) concentrations in leaves of F. americana measured 1 week and 1 month after trees were injected with different injection systems.
Effects of Injection Method on Efficacy
Mean canopy thinning at the Culver Academy site (Figure 1) was greatly affected by treatment (F(3,64) = 29.21, P < 0.0001) and year of evaluation (F(3,64) = 5.66, P = 0.0017). During the first year of the study, canopy thinning was not significantly different among treatments (P = 0.87). However, canopy thinning in untreated trees increased significantly more than in treated trees 1 and 2 years after injection (P = 0.004), but there were no differences in canopy thinning among trees treated with insecticides (P > 0.05). In 2019, 3 years after injection, mean canopy thinning increased in trees treated with the enTREE system to 32%, whereas it increased to 12% in trees treated with the QC and 6% with the Tree I.V. systems, respectively. Final mean canopy thinning of trees treated with the enTREE system was significantly higher than trees treated with the Tree I.V. system (P = 0.0087) but not significantly different from trees treated with the QC system (P = 0.122). All treatments still had less canopy thinning than the untreated control (P < 0.001).
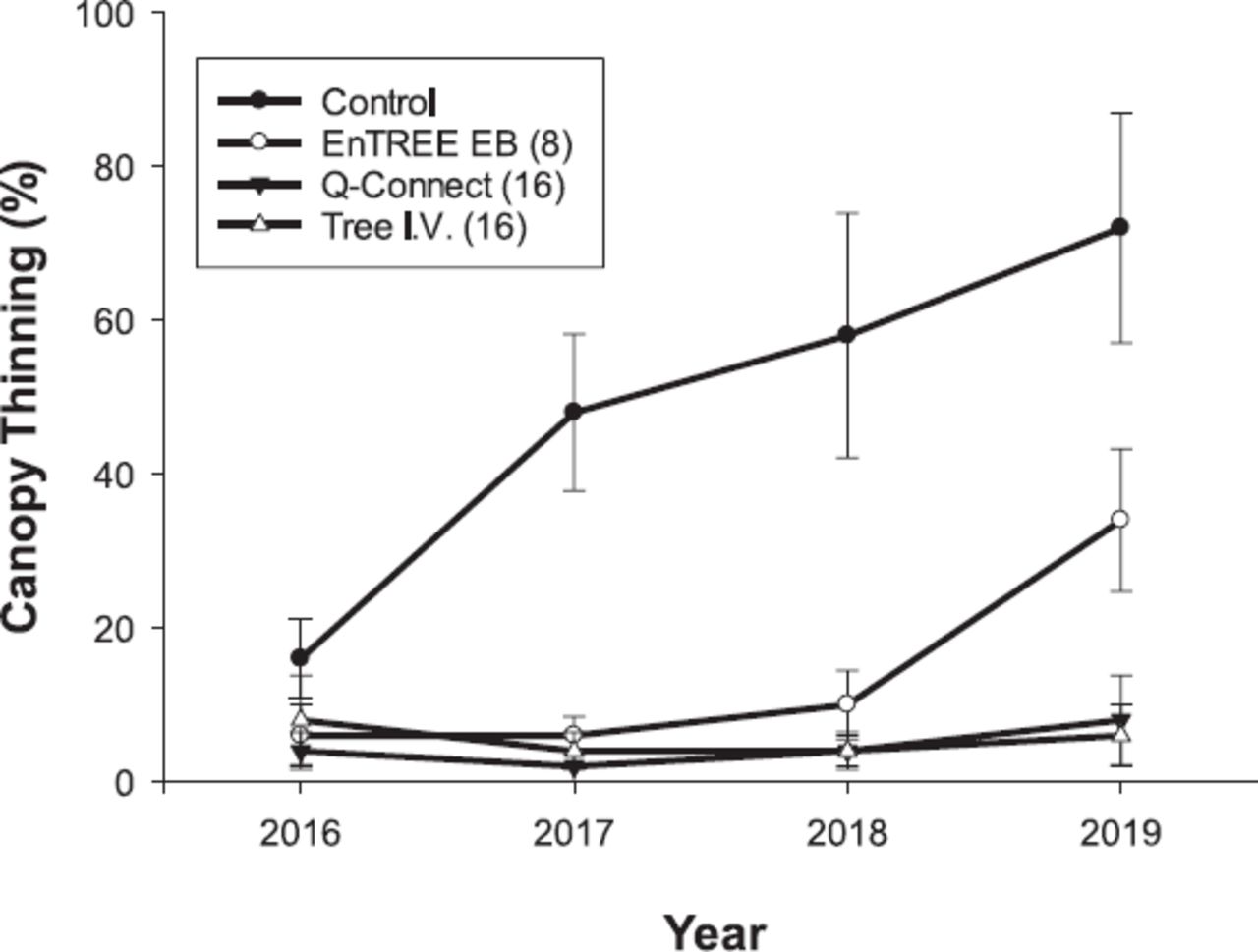
Canopy thinning of white ash trees caused by emerald ash borer when treated with 4% emamectin benzoate in 2016 June with 3 different injection devices and an untreated control in Culver, IN, USA (see text for explanation).
Effects of Application Timing on Duration of Efficacy
Mean canopy thinning at the Eagle Creek Park site (Figure 2) was greatly affected by treatment (F(2,208) = 282.48, P < 0.0001), time (F(7,208) = 27.79, P < 0.0001), and the treatment × time interaction (F(14,208) = 10.24, P < 0.0001). At the start of the study (2013), there were no significant differences in canopy thinning among treatments (P > 0.05). In 2014, 1 year after injection, trees treated in the spring had less thinning than the untreated control (P = 0.046), whereas thinning of trees treated in the fall did not differ from the control (P = 0.731). By 2015, significantly more canopy thinning was detected in the untreated trees than in those treated in spring or fall (P < 0.0001). However, at this time, trees treated in the fall had significantly more canopy thinning than those treated in the spring (P = 0.021). Canopy thinning of trees treated in the fall and the spring peaked in 2016 and then declined annually after the 2016 retreatment until 2020 when canopy thinning began to rise again in both treatments.
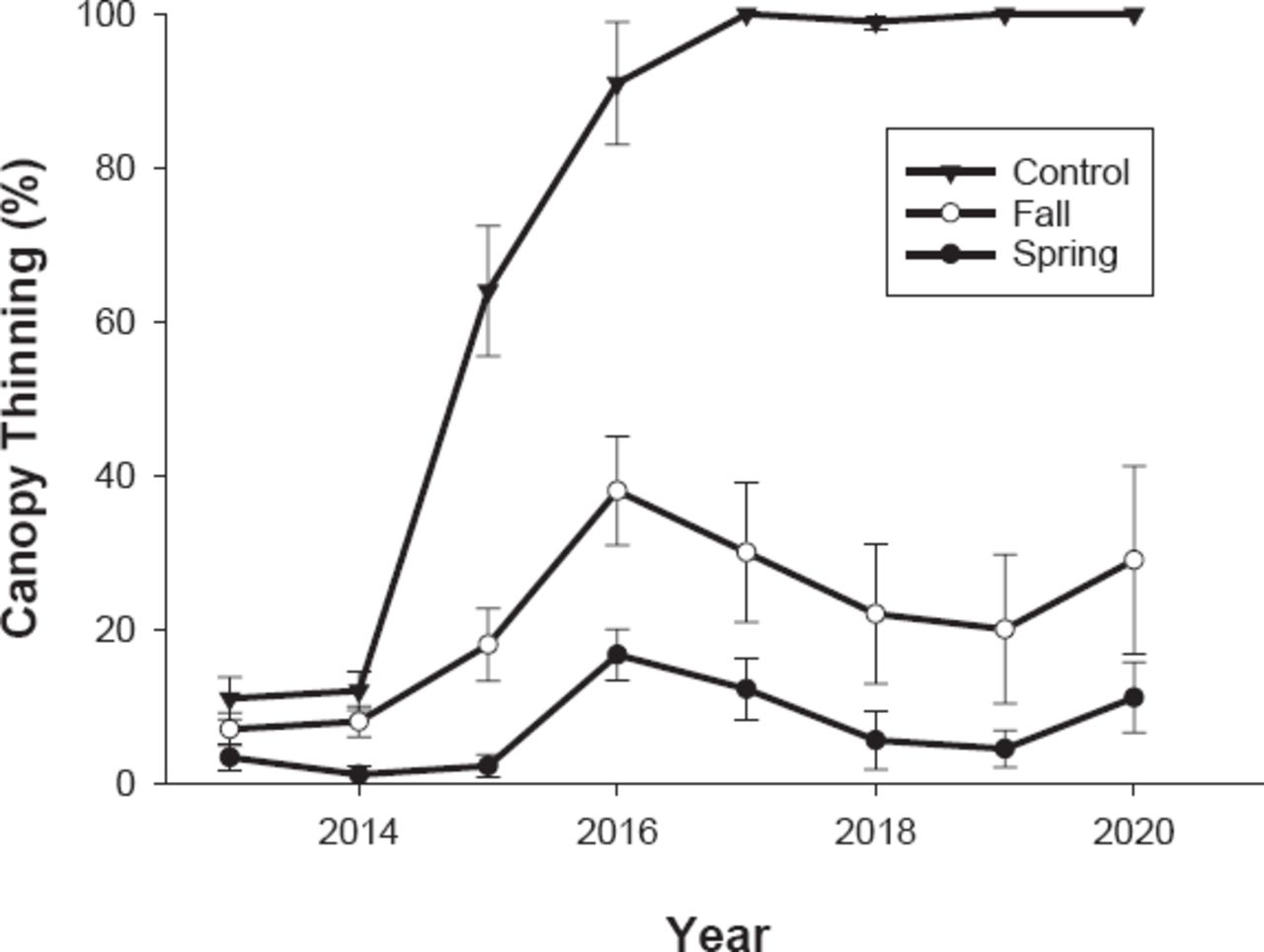
Canopy thinning of white ash trees caused by emerald ash borer treated with 4% emamectin benzoate in spring and fall of 2013 and 2016 in Eagle Creek Park, Indianapolis, IN, USA, with the Tree I.V. injection system.
DISCUSSION
Our study clearly demonstrates that EB can be successfully delivered to the canopy of ash trees by the 3 injection systems we evaluated. Even when the dose per tree was held constant, the ability of each system to protect a tree from EAB varied based on how the characteristics of each injection system impacted the uptake of insecticide into the canopy. A single application of insecticide in the spring was more efficacious than if applied in the fall, suggesting that the ability of EB to protect ash trees was influenced by the seasonal phenology and population dynamics of EAB. Applying EB in the spring when adults and larvae were actively feeding allows trees to avoid the injury that would have occurred in summer prior to fall injection of trees. Arborists need to consider these factors when developing strategies to deploy EB treatments for EAB.
The number of injection ports required for each system had the most significant impact on the capacity to deliver insecticides to the canopy and protect trees. In fact, the injection systems with 16 ports/m of trunk circumference (Tree I.V. and QC) had a more uniform distribution of toxicant to leaves and a longer capacity to protect trees than the system with half the number of ports. The more uniform EB distribution with systems employing more ports is consistent with the straight line patterns of uptake associated with the sectorial vascular system of ash trees (Chaney 1986). It is also supported by studies of radiolabeled imidacloprid, which showed that more product was taken up into the leaves of ash tree branches that were in the same vertical plane of the injection site (Tanis et al. 2012). The 16-port QC injection system was similar to the 8-port enTREE system in several ways, including the same EB formulation, the same diameter injection port, and the absence of plastic plugs. The other 16-port system, Tree I.V., used a different EB formulation, larger injection port, and proprietary port plugs to prevent leakage. Despite this, no differences were found in the concentration of EB detected in leaves or the proportion of leaves with insecticide between trees injected with the Tree I.V. and QC 16-port systems. This lack of difference between these 2 injection systems suggests that the factors such as diameter of the holes drilled, presence or absence of plastic plugs, and EB formulation had no significant effect on delivery of the active ingredient into the leaves. Both systems delivered over 1,000 ppb of EB into the leaf tissue, a level of detection known to kill adults in laboratory bioassays and associated with densities of larvae near 0 in replicated studies (McCullough et al. 2011).
With only 64% of leaf samples infused with EB a month after being treated with the enTREE system, it seems likely that the canopy became a mosaic of toxic and nontoxic leaves. Given that adult female EAB feed on leaves for 1 to 2 weeks before laying eggs, it is quite likely that many adult females would have encountered a toxic leaf before laying eggs during 2016 and 2017. This could explain why, despite a more uneven distribution and lower concentration of product in the leaf canopy of trees treated with the enTREE system, each injection system provided effective control compared to the untreated trees in 2016 and 2017. This explanation is consistent with mechanisms used to explain how area-wide pest management programs can slow EAB population growth and reduce ash mortality when only a subset of the trees are treated (McCullough and Mercader 2012; Sadof et al. 2021).
Despite 2 years of no increased canopy thinning, after the third season of EAB activity in 2018, trees treated with the enTREE system experienced numerically more thinning than trees treated with the other 2 injection systems. By 2019, after a fourth season of EAB, this difference in canopy thinning became significant. The reduced protection of trees treated with the enTREE system could be linked to the sectorial distribution of EB through trunk and branch xylem. While larvae feeding within the plane of insecticide distribution were likely killed by EB, larvae outside this plane could survive, feed on portions of the vascular system, and reduce the capacity of branches to support the production of foliage. If the trees treated with the enTREE system received a second application in the spring of 2018, as recommended by the product label, it is possible that the trees would have avoided injury from EAB during the 2018 and 2019 growing seasons.
Our long-term study with the Tree I.V. system at Eagle Creek Park suggests that the capacity of EB to protect a tree from EAB can also be influenced by the timing of the application. This study was initiated early in the infestation cycle when the average canopy thinning was 8.2%. Trees injected in the spring are protected since females are killed before they are able to lay eggs and because young larvae are killed before they can do much injury to the phloem. Trees injected in the fall experienced EAB injury throughout the summer prior to treatment. Recent field studies of EAB phenology indicate that while the majority of EAB larvae (90%) were actively feeding third to fourth instars until the end of September (< 1,700 DD10), some larvae (32%) actively fed until the end of October (Jones et al. 2020). The elevated thinning on fall-treated trees may have been avoided if the second fall applications were made a year earlier in 2015. Other workers have also found that soil drenches of imidacloprid were more effective when applied in the spring than fall (Smitley et al. 2015). Although fall treatments provide arborists a longer window in which to apply insecticides to EAB-infested trees, spring applications are likely to be more effective because they are more closely aligned with the phenology of actively feeding adults and larvae.
Differences in the longevity of control afforded by each application of insecticide may be explained by changes in the population dynamics of EAB over time. Studies of tree mortality and the number of beetles emerging from a stand of infested trees show that by the time 90% of trees had died, most of the beetles (90%) that would be produced by the standing ash trees had already emerged (Steiner et al. 2019). Thus, we assume that in 2015, when canopy thinning in the untreated controls had risen to 64%, the population of adult beetles in the forest was most likely nearing its peak. Based on the increase in canopy thinning between 2015 and 2016, beetle abundance at this time was likely sufficient to damage trees in 2015 on both treated and untreated trees. Thus, trees treated in 2013 were afforded only 2 years of protection. In contrast, when trees were treated again in 2016, spring and fall treatments stopped further injury for 3 years through 2019. In 2016, after 91% of the canopy was missing from the untreated control trees, EAB populations were probably past their peak because most of the available phloem to support beetle development in the forest had already been consumed. The increase in canopy thinning in 2020 reflected successful colonization of treated trees after concentrations of toxicant had dissipated.
CONCLUSION
Our study has important implications for arborists. Municipal arborists who seek to optimize the benefits of their protection investment to the entire urban forest will be able to inject trees once every 3 years, provided that they apply the insecticide in the spring and use systems with more injection ports. Control programs that use systems with fewer ports like enTREE or fall applications should be on a 2-year cycle. Residential arborists, or those whose client base has little tolerance for treatment failure, may choose to treat on a 2-year cycle, especially when local populations of EAB approach their peak. After the bulk of EAB has emerged from trees infested during the initial wave of EAB invasion, residential arborists may lengthen the application interval to 3 years within the context of a broader scouting-based plant health care program.
ACKNOWLEDGMENTS
We thank Paul Pinco and William Kincius of the Indianapolis Parks Department, and David Blalock of Culver Academy for permission to conduct research on their properties. We thank Bruce Cooper and Amber Hopf Jannasch at the Purdue University Bindley Bioscience Center for assistance with the pesticide residue analysis. We also thank S. Stack and D. Peterson for technical assistance. Insecticides and application equipment were provided by Rainbow Treecare, Arborjet, Inc., and Brandt Corporation. This project was funded in part by funds supplied by Rotam North America, Arborjet, Inc., and USDA NIFA Hatch Project Managing Emerald Ash Borer in Urban and Natural Forests IND 011478 to CSS and MDG.
Footnotes
Conflicts of Interest:
The authors reported no conflicts of interest.
- © 2022, International Society of Arboriculture. All rights reserved.
LITERATURE CITED
In this issue








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.