
Abstract
Two experiments investigated factors influencing root penetration of polyvinyl chloride (PVC) pipes. Eucalyptus leucoxylon, Allocasuarina littoralis, Lophostemon confertus, Callistemon salignus, Acer palmatum, and Pyrus calleryana seedlings were grown in containers containing 150-mm lengths of sealed 75-mm PVC stormwater pipe with cracks 0.04 mm, 0.66 mm, or 1.48 mm wide on their upper surface. The buried pipes contained water, water and stormwater, soil, or soil and stormwater. There were six replicates and 432 plants. There was no significant difference in the mass of roots entering the pipes for the two larger crack widths with 70% of pipes penetrated and strong growth inside the pipes. While the roots of all species penetrated cracks greater than 0.66 mm, only roots of C. salignus, E. leucoxylon, and L. confertus penetrated 0.04 mm cracks. Roots penetrated 50 to 60% of pipes containing soil, or soil and stormwater, and 40% of pipes containing water, or water and stormwater were penetrated. The plants with roots penetrating pipes containing water and stormwater grew tallest. No roots penetrated the welded caps of the stormwater pipes. A second experiment using E. leucoxylon, Melaleuca ericifolia, Ficus macrophylla, A. littoralis, and Salix fragilis investigated root penetration of different sized holes in polycarbonate plates. The plates, installed in containers with growing medium above and below, had either 2 × 4 mm holes, 8 × 2 mm holes, 127 × 0.5 mm holes, or a mixture of holes (1 × 4 mm, 2 × 2 mm and 32 × 0.5 mm holes), total pore area in all being 25.14 mm2. Below the plates, the growing medium was capillary irrigated with stormwater or water. All species grew through 0.5-mm holes and had strong root growth below the plates. When irrigated with stormwater, all species were taller and had greater biomass, and most species had a greater root mass below the plates. In general and regardless of hole size, the more holes in the plates, the more roots penetrated them. Properly installed PVC pipes are impenetrable, but the width and number of openings in a pipe influence the capacity for penetration and subsequent root growth so protocols minimizing damage to pipes should be enforced. Since species have different capacities for penetrating stormwater pipes, appropriate species selection for urban environments where damaged pipes may occur could reduce incidences of pipe damage.
INTRODUCTION
Trees are important parts of urban infrastructure, providing environmental, human health, and social benefits (Lohr et al. 2004). Trees evolved in forests and woodlands, not in urban soils in narrow verges between foot paths and roads where they share rooting space with sewerage and stormwater pipes and other utilities. These unnatural rooting spaces lead to compromises in tree health and conflicts with infrastructure.
Disturbed and often compacted soils have reduced oxygen diffusion and drainage, limiting tree root growth (Kozlowski 1992; Craul 1994; Lichter and Lindsey 1994; Bradshaw et al. 1995). Backfill around pipes is often less compacted than the surrounding soil, making it suitable for roots, which can then penetrate pipes if there are cracks larger than the root tips (Rolf et al. 1995; Stal and Rolf 1998; Pohls 2001). Roots require pore spaces larger than the growing tip for elongation, although they can push through pores, enlarging them in friable, uncompacted soil (Roberts et al. 2006; Urban 2008). Tree roots can grow into cracked pipes, creating costly problems for local governments (Lesser 2001; Dodge and Geiger 2003; Lohr et al. 2004). Trees are often removed if their roots cause even minor infrastructure damage (Hannah and Yau 1990), which usually involves the removal of a valuable, mature tree that is then replaced with a younger specimen.
Moisture condenses on the outside of clay pipes (Roberts et al. 2006), and opportunistic roots attracted to the moist and fertile environment which exists at leaks can enter the pipes at cracks rather than causing cracks (Ridgers et al. 2006). There is greater condensation around clay or concrete than around polyvinyl chloride (PVC) pipes (McPherson and Peper 1994). Most roots enter clay and concrete pipes through breaks, loose joints, or failed rubber gaskets, and smaller pipes (diameter 22.5 to 40 cm) are most often penetrated, especially in sandy soils (Watson et al. 2014). Water mains have sufficient water pressure to prevent root growth, but this is not the case for sewers and drains that rely on gravity to move their contents (Roberts et al. 2006).
Roots will grow along moisture gradients created by leaky pipes or by condensation around the pipes (Coder 1998; Randrup et al. 2001; Roberts et al. 2006). Once they have penetrated pipes, roots grow rapidly, causing blockages, but over time the blockage may cause anaerobic conditions, and the roots die back (Ridgers et al. 2006). Damage may occur on shrinkable clays, when roots grow under pipes and increase in diameter, if pipes are encircled by roots, or if strong winds cause sufficient root movement to break pipes (Watson et al. 2014). When pipes are warmer than the surrounding soil, fine root activity is greater (Kurpaska et al. 2005; Kawasaki et al. 2014).
Older sewers were made of vitrified clay, brick, and concrete, while more modern systems are made of plastic, ductile iron, steel, and reinforced concrete, which have adequate compressive strength and sufficient tensile strength to reduce cracking and damage. Between the world wars in England, clay packed joints and cement seals dried over time and cracked (Rolf et al. 1995; Randrup et al. 2001). In the 1930s, joints of drainage pipes contained copper wire to prevent root growth, but these eventually failed (Roberts et al. 2006). Clay and concrete pipes without rubber joint gaskets are likely to have faulty joints (McPherson and Peper 1994; Rolf et al. 1995). In Sweden, most intrusions occurred in pipes of 225 to 400 mm diameter with yarn and cement sealed joints laid before 1950. The rubber seals of the 1960s were more effective, and the roots growing in pipes from that era are probably due to shoddy work.
In Melbourne, the most common occurrences of roots in pipes were in thirty- to sixty-year-old clay pipes joined with rubber rings located at a depth of less than two meters (Pohls 2001). These seals expand and break, and joints were the most common sites of root intrusion (Coder 1998; Stal and Rolf 1998). PVC plastic and fiberglass pipes that are laid correctly with welded joints are rarely invaded by roots if they are installed according to correct standards on a compacted bed of gravel (McPherson and Peper 1994; Coder 1998; Randrup et al. 2001).
While roots rarely break pipes, prevailing winds affecting the root plate may place stress on pipes on the windward side of the tree. Roots growing on the windward side of the tree are under tension and refer this tension to pipes lying over them, which may crack on their upper surface, especially if the roots are of a large diameter. Pipes directly under the trunk and pipes on the leeward side of the tree are not at risk of damage (Roberts et al. 2006).
Different species of trees are implicated in pipe damage in different parts of the world. In Melbourne, the roots of commonly planted eucalypts and Melaleuca species were most frequently found in pipes (Pohls 2001), while in Sweden, willow (Salix) and poplar (Populus) were the main cause of damage (Rolf et al. 1995; Stal and Rolf 1998). Pipes containing stagnant water are not conducive to root growth due to a lack of oxygen, but willow and swamp cypress (Taxodium) can grow in these conditions, and pipes that have water flow twenty-four hours a day, are more likely to have root growth than drains that carry water occasionally (Roberts et al. 2006).
Reduced root growth can be achieved by annual cutting, flooding pipes with boiling water after cutting roots, or using copper sulphate, which is only taken up a short distance into the root before the root dies, preventing further absorption without killing the tree (Rolf et al. 1995; Randrup et al. 2001). Exposure to copper sulphate requires days to weeks to kill roots, which is often impractical. Chemicals such as dichlobenil, oryzalin, and trifluralin control root growth, but affect water quality (McPherson and Peper 1994; Pohls 2001). Metham (methylcarbamadithioic acid) has been used for control but is toxic to sewage treatment nitrifiers and was replaced by less toxic but more costly glufosinate. Herbicides have been used to control root growth for three to five years, but many have faced environmental bans in Europe (Roberts et al. 2006).
Two experiments investigated the penetration of PVC pipes by the roots of different tree species. The first experiment investigated the penetration of PVC pipes by roots of six commonly planted street trees. The pipes had cracks of different widths cut into them and contained potable water, stormwater, sandy soil, and soil plus stormwater. The research aimed to determine which of the species were most likely to penetrate the cracks, how crack size influenced root penetration, and whether the content of the pipes affected penetration and subsequent root growth. The second investigated the capacity of the roots of four tree species to grow through holes of different sizes drilled into polycarbonate laser-cut discs. Below the discs, media were irrigated with potable water or stormwater, and root and canopy growth were measured. This experiment investigated whether the size and number of holes in a pipe influenced root penetration. In both experiments, plants were irrigated with water and stormwater to examine whether root penetration and proliferation were greater in pipes containing stormwater that contained higher levels of some nutrients.
MATERIALS AND METHODS
Experiment 1
Six species commonly grown as urban trees in Melbourne, Australia were grown at the University of Melbourne, Burnley College, 500 Yarra Blvd, Richmond: Acer palmatum Thunb (Japanese maple), Allocasuarina littoralis (Salisb.) L.A.S. Johnson (black she-oak), Callistemon salignus (Sm.) DC (pink tips bottlebrush), Eucalyptus leucoxylon F. Muell. Subsp. megalocarpa Boland (yellow gum), Lophostemon confertus (R. Br.) P.G. Wilson and Waterhouse (brushbox), and Pyrus calleryana Decne (callery pear). Seedlings of all species were 350 +/− 100 mm tall at planting and were grown for ten months between late-January and mid-December. Plants were grown in soil purchased from Burdetts Nursery, 615 McClelland Drive, Langwarrin, which consisted of 60% whitewashed sand, 25% screened soil, 5% chicken manure, and 10% organic compost by volume. The soil had a drainage rate of 6 mm/hr and a wettability rate of 60 mm/min (Australian Standard 4419-2003 using compacted soil and three replicates). Particle size of the soil was also determined according to Australian Standard 4419-2003 USGA sieve 1993 (Table 1). Soil pH and electricity conductivity were measured in 1:5 extracts, and the data sheet provided by the supplier showed the pH to be moderately acidic (6.0 to 6.6) while the total salts were low (electrical conductivity < 0.6 dS/cm). Plants were irrigated by overhead sprinklers and fertilized with 12 mL Maxicrop diluted in 4 L of water in May and then irrigated with 17.8 mL Peters liquid fertilizer (15N:11P:25K) diluted in 37.8 L of water in mid-August and mid-October.
Particle size analysis of the soil used to grow plants and inserted into pipes.
PVC stormwater pipe (75 mm diameter) was cut into 150-mm lengths and had cracks of 100-mm length cut into them. The narrowest cracks were made by squeezing the pipes in a vice and cutting with a Stanley knife. These cracks averaged less than 0.04 mm wide, measured with an engineer’s feeler gauge, which measures the gap between two solid components, and the gap width was the average for ten pipes. The medium cracks were cut using a Dremel saw with a fine cutting wheel and averaged 0.66 mm wide, and the large cracks were cut using a circular saw and averaged 1.48 mm wide. Caps were welded onto the ends of the pipes using Plaskeen solvent cement containing methyl ethyl ketone, as recommended by the manufacturer.
The experiment was a randomized complete block design. The treatments were a combination of three factors (crack, media, species), each having multiple levels. There were three widths (small, medium, large), four different media (water, stormwater, soil, soil + stormwater), and six species. There were six replicates for each of these 72 treatments and a total of 432 plants. Positions of containers in the six blocks were chosen by using a random number generator for each block.
Soil was inserted into the already cracked pipes with one cap glued on and the crack taped over with duct tape, and then 450 mL of water was added to a quarter of the pipes before welding the second cap. Similarly, 450 mL of stormwater was added to another quarter of the pipes. Another quarter of the pipes were filled with the same dry sandy soil used to grow the plants. The remaining pipes were filled with 180 mL of stormwater, which saturated the soil.
The stormwater synthesized for this trial was based on synthetic runoff to bioretention systems (Davis et al. 2006) (Table 2). The concentration of the dissolved solids was varied by replacing calcium chloride with sodium chloride and magnesium chloride to more closely reflect the composition of stormwater in Melbourne (Denman 2009) with a pH of 6.4.
Chemical composition (mg/L) of stormwater solution (Denman 2009).
Seedlings were planted in 250 mm containers with 10 mm of fine composted pine bark placed at the base of the containers to prevent the sandy loam from washing from the pots. The prepared pipes were placed crack-side up in the base of the container. Soil was then packed in around the pipe to keep it crack-side up. Turf blend was placed over the pipe, and the seedlings were planted in turf blend before being placed in their positions and watered (Figure 1).
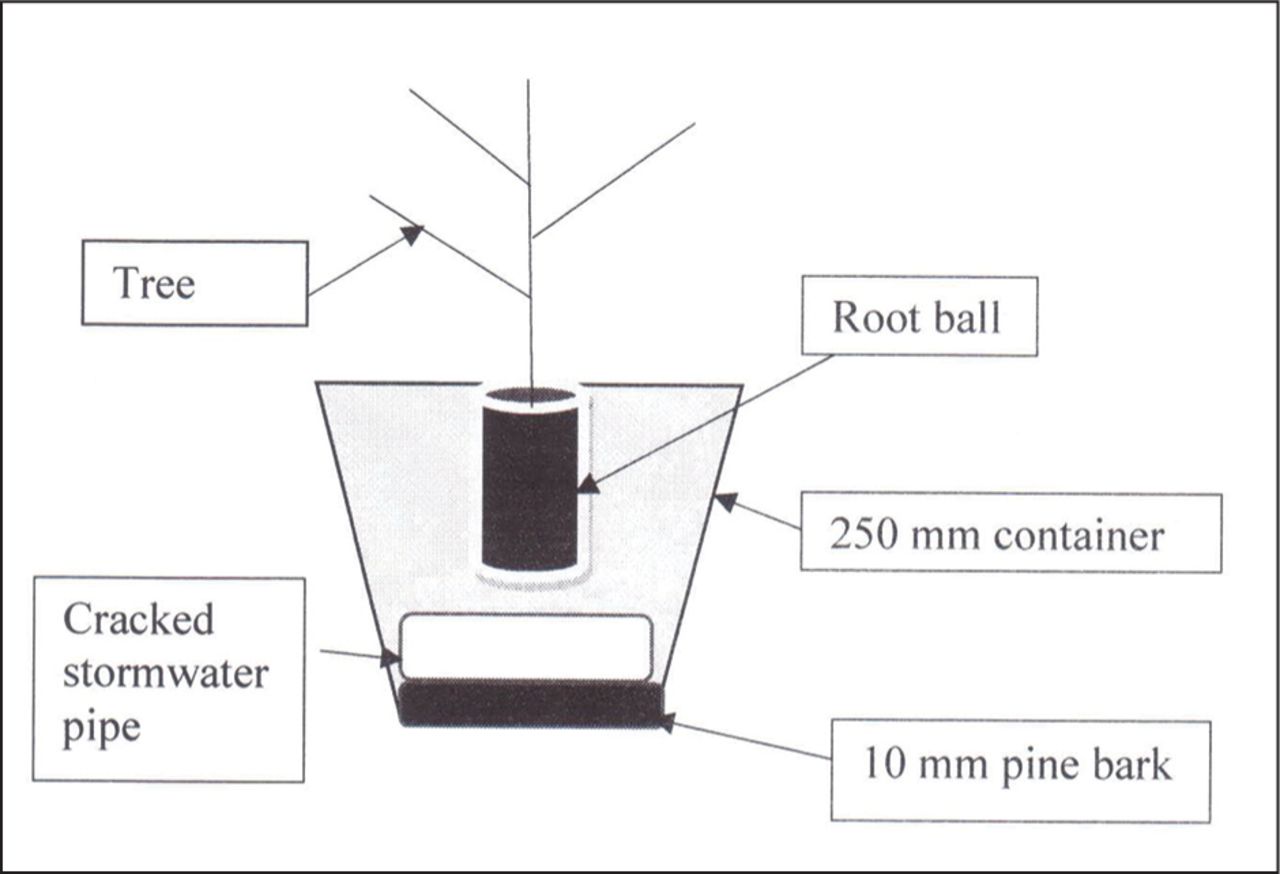
The planting design for experiment 1 showing the seedling positioned above a PVC pipe with a crack on the upper surface.
Height and canopy spread (north-south and east-west) were measured three times over the course of the experiment (February, June, and November). On completion of the experiment, stems were cut at soil level, and shoots and roots were placed in separate bags and dried prior to the determination of oven dry weight (80° C for 48 hours).To harvest the roots, the pots were gently upturned, making sure the roots growing into the pipes were not disturbed. The roots were cut where they entered the pipes, and the pipes were then removed for later extraction of roots. The roots not in the pipes were then gently shaken and washed to remove the sandy loam and any medium. The pipes were removed from the containers, the cap from each pipe was removed, and the contents, if any, were placed in numbered bags. The roots from the soil and soil and stormwater filled pipes were gently shaken and then washed as above.
Data were analyzed using Minitab 16. The normal probability plots of distribution of residuals were justified using the ANOVA General Linear Model (GLM) if normally distributed or otherwise for using nonparametric tests. For ANOVAs, P-values of less than 0.05 were considered significant, and the least significant difference (LSD) was calculated for each pair of means. When analyzing the data for the mass of roots in pipes for effects of crack width, species, and contents of fill, data were analyzed for the 210 plants that had roots penetrate pipes and separately for all 432 plants. If the plot of distribution of residuals was not an approximation to a straight line, the distribution of residuals was transformed to log10. If this distribution of residuals did not result in a normal distribution, the data were analyzed using nonparametric Kruskal-Wallace or Mann-Whitney Tests. The Mann-Whitney Tests were applied when the P-value for the Kruskal-Wallis Test was below 0.05.
Experiment 2
Four urban trees species grown in Melbourne, Australia were grown in a greenhouse at University of Melbourne, Burnley College for 16 weeks from July to October: Ficus macrophylla Desf. Ex Pers. (Moreton Bay fig), Melaleuca ericifolia Smith (swamp paperbark), Salix fragilis L. (crack willow), and Eucalyptus leucoxylon F. Muell. Subsp. megalocarpa Boland (yellow gum). Trees were planted in olive pots (square tapered containers 90 × 90 mm × 180 mm deep) placed on capillary matting along two benches of the greenhouse (Figure 2).
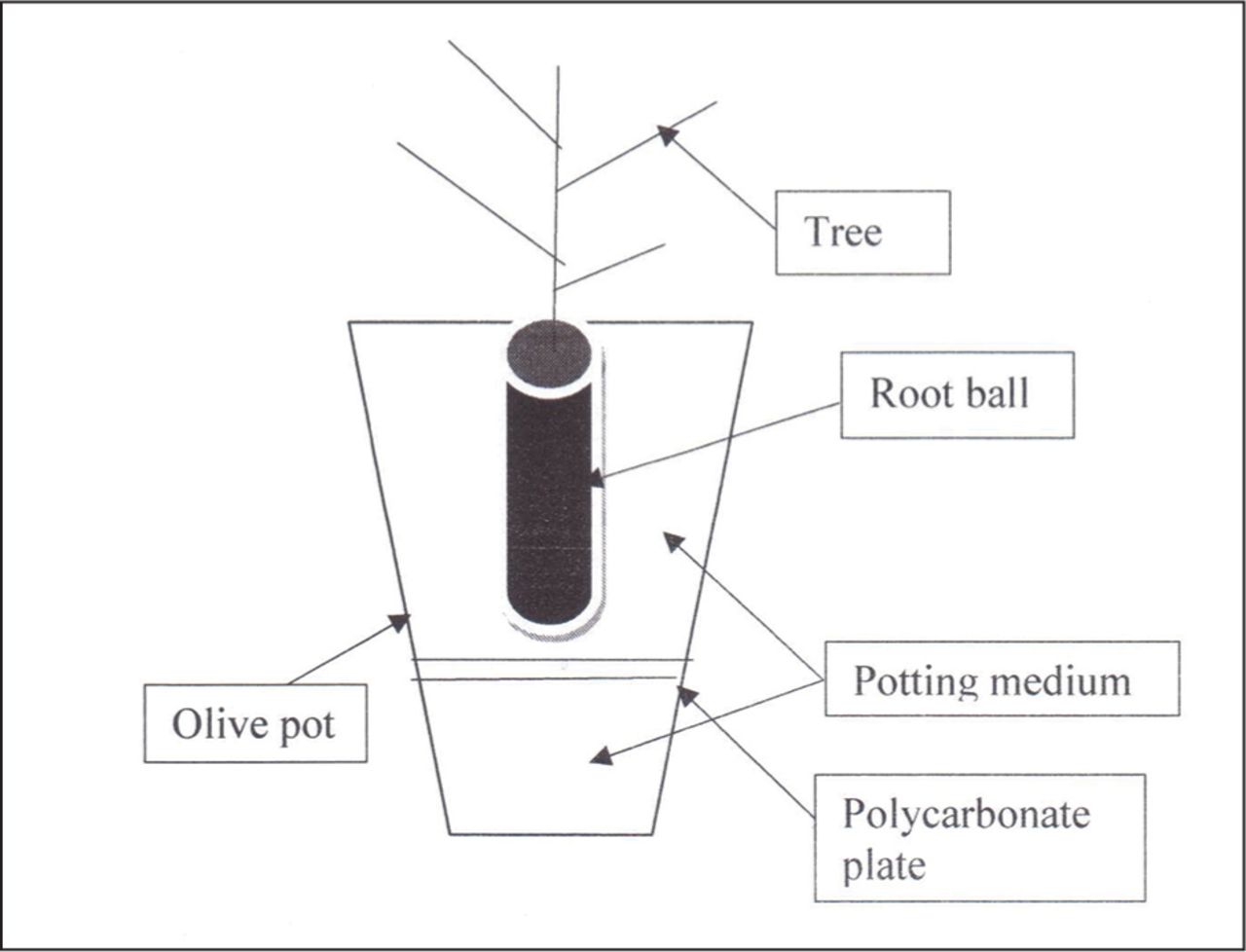
The planting design for experiment 2 showing the seedling positioned above a polycarbonate plate with different configurations of hole sizes and numbers of holes.
Polycarbonate plates were laser cut, and four series of holes were drilled in the middle of the plate. One plate had 2 × 4 mm holes, another had 8 × 2 mm holes, one had 127 × 0.5 mm holes, and one had a mix of holes (1 × 4 mm, 2 × 2 mm, and 32 × 0.5 mm holes). These arrangements gave a total aperture area of 12.56 mm2 for every plate. There was a control with no plate, giving five treatments. There were two irrigation regimes (water, stormwater) and four species with six replicates. This gave a total of 240 plants (5 types of plate × 4 species × 2 irrigation regimes × 6 replicates). There were six blocks of 20 plants on each side of the greenhouse, and plants were allocated randomly within each block. The six blocks of 120 plants on one side of the glasshouse were irrigated with tap water, while those on the other were irrigated with stormwater (Table 2).
Trees were grown in a soilless medium (pine bark without fertilizer) so that differences in growth from water or stormwater (Table 2) irrigation could be determined. Plants were planted into olive pots with 225 mL of potting medium placed in the base of the pot. The plates were jammed horizontally on the potting medium layer (Figure 2) but were not sealed so that roots below the plate could be harvested. A small amount of potting medium covered the plate, and the plants were then potted up.
On completion of the experiment, the stems were cut at soil level, and shoots and roots were placed in separate bags and dried prior to the determination of oven dry weight (80° C for 48 hours). To harvest the roots, the containers were gently upturned, ensuring roots growing through the holes in the plates were not disturbed. The roots were cut immediately above the plate, and the number of roots passing through each hole noted. For controls, the medium was sliced at the same level—immediately above the plate. Roots were gently shaken and washed free of medium.
Data were analyzed as for experiment 1 using the statistical package Minitab 16. Again, Kruskal-Wallis and Mann-Whitney Tests were applied as necessary. As plants irrigated by water and plants irrigated by stormwater were grown on different sides of the glasshouse, each irrigation method was analyzed separately.
RESULTS
For some experiments, the plots of the distribution of residuals meant that data were analyzed using nonparametric tests. Only statistically significant (P < 0.05) results are reported.
Experiment 1
There were significantly fewer root penetrations of pipes with the narrowest crack width compared to both the medium and larger crack widths, which did not differ from each other (Figure 3). Only five (3.5%) roots penetrated the narrowest crack, compared to 104 (72.2%) and 101 (70.1%) roots for the medium and larger cracks, respectively.
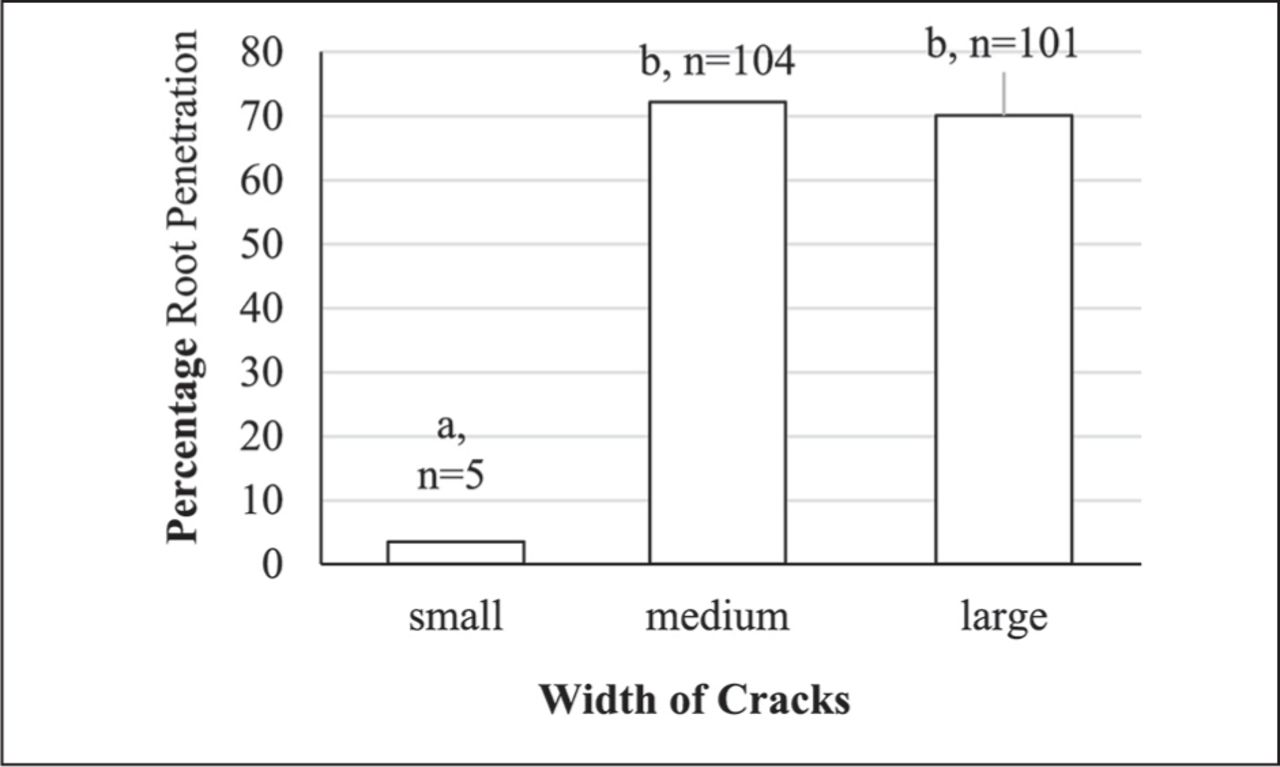
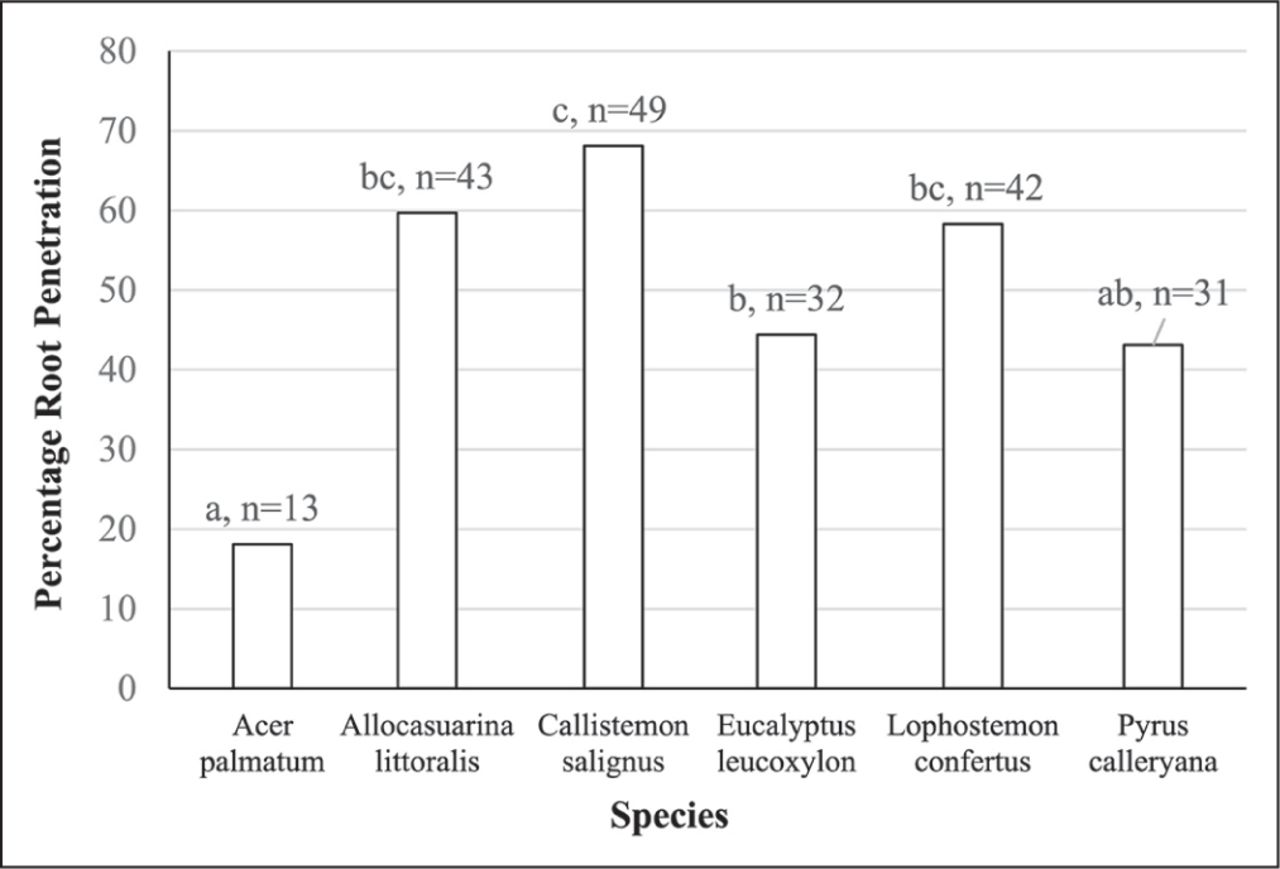
Root penetration of pipes with different width cracks (%). The numbers at the top of columns are the number of roots that penetrated pipes, and the height of the column provides the percentage of root penetrations for the total number of plants (N = 144). Means with a different letter are significantly different (P < 0.001).
There was also a significant difference in the root mass found in pipes with the narrowest crack width compared with the medium and larger crack widths (P < 0.001), which again did not differ from each other (P = 0.9501) (Table 3).
Mann-Whitney Tests comparing of mass of roots penetrating the pipe for different crack widths for all plants (N = 144).
It was anticipated that species would influence crack penetration. A. palmatum had the fewest penetrating roots, followed by P. calleryana and E. leucoxylon (Figure 4). Root penetrations by A. palmatum and P. calleryana were not significantly different from each other, but A. palmatum differed significantly from A. littoralis, E. leucoxylon, P. calleryana, and L. confertus.

Root penetration of pipes by different species (%). Means with a different letter are significantly different (P < 0.001).
C. salignus and P. calleryana had the greatest root mass penetrations, and while A. littoralis and L. confertus were similar in mass, the root lengths of A. littoralis were shorter (Table 4). The root mass penetrating pipes for A. palmatum was not significantly different from E. leucoxylon, but they were significantly different from the other four species (Table 4).
Mann-Whitney Tests comparing mass of roots penetrating pipe for all plants of species that penetrated the pipes.
There were no significant differences in the number of roots penetrating pipes filled with potable water or stormwater. However, the root mass in pipes filled with soil (0.03 g) was significantly higher than those filled with stormwater (P = 0.035) and higher for water-filled pipes (P = 0.063), both being 0.001 g. There were no significant differences in the heights of trees, which ranged from 72 to 78 cm, and masses, which ranged from 44 to 50 g, for any of the treatments or for whether they penetrated pipes.
Experiment 2
Table 5 compares the root mass below the plates for different hole configurations when irrigated with water. There was a significant difference for controls compared to all hole configurations (P < 0.05), but no difference between plates. The plates limited root growth below them.
Mann-Whitney Tests comparing median root mass below different configurations of holes in plates irrigated by water (N = 24 except for 2 mm holes where N = 22).
Table 6 compares the mass of roots below the plates for different species irrigated with water. The pairing of F. macrophylla and S. fragilis was not significantly different (P = 0.367), but all other pairings were significantly different (P < 0.05).
Mann-Whitney Tests comparing median root mass below plates for different species irrigated by water (N = 29 for E. leucoxylon and S. fragilis, and N = 30 for M. ericifolia and F. macrophylla).
The mean mass of roots below the plate for plants irrigated by stormwater was significantly greater for controls than for containers with plates (Figure 5). Plates with holes of 0.5 mm diameter had significantly greater root mass below the plates than plates with holes of 4 mm diameter.

Mean mass of roots for all species below the plate for plants irrigated by stormwater against hole configuration. Means with a different letter are significantly different (P < 0.05).
The mean mass of roots below the plate for plants irrigated with stormwater was significantly greater for S. fragilis than the other three species (Figure 6), and F. macrophylla had a significantly greater root mass below the plates than E. leucoxylon.

Mean mass for roots below the plate for all hole configurations for different species irrigated by stormwater. Means with a different letter are significantly different (P < 0.05).
The P-values from the Kruskal-Wallis Test for mass of roots below the plates for species irrigated by water were significantly different for both hole configuration (P = 0.014) and species (P < 0.001) (Table 7).
Kruskal-Wallis test for roots below the plate for species irrigated by water (N = 120).
A plot of the distribution of residuals for log10 (mass of roots below plate) for plants irrigated with stormwater was normally distributed, and these data were subjected to an ANOVA GLM (Table 8). There were significant differences for species (P < 0.001) and hole configuration (P = 0.004), but not for the interaction of hole configuration with species (P = 0.589).
General linear model for log10 (mass of roots below plate) for plants irrigated by stormwater (N = 120).
For plants irrigated with water or stormwater, there were differences in total plant biomass for different species (Figure 7). For plants irrigated with water, differences in hole configurations were not significant, but they were for plants irrigated with stormwater (P < 0.001). The interaction of species and hole configuration was not significant. Plants irrigated with stormwater had consistently greater mass than those irrigated with water (Figure 7).

Mean total plant biomass of species irrigated by water (white columns) and stormwater (grey columns). Vertical bars show Least Significant Difference (P < 0.05).
The control plants had a greater mass than those with perforated plates, and plants irrigated with stormwater were taller and heavier than those irrigated with water (Figure 8).

Mean biomass of plants for different hole configurations irrigated with water (white columns) and stormwater (grey columns). Vertical bars show Least Significant Difference (P < 0.05).
The ANOVA GLM for percentage increase in height of plants against species irrigated by water and stormwater (P < 0.001) was significantly different (Table 9). The P-value for percentage increase in height of plants against hole configuration (P = 0.012) and for the interaction of hole configuration with species (P = 0.038) were significantly different in both cases.
Percentage increase in height of plants irrigated by water or stormwater (N = 120, separate ANOVA GLMs).
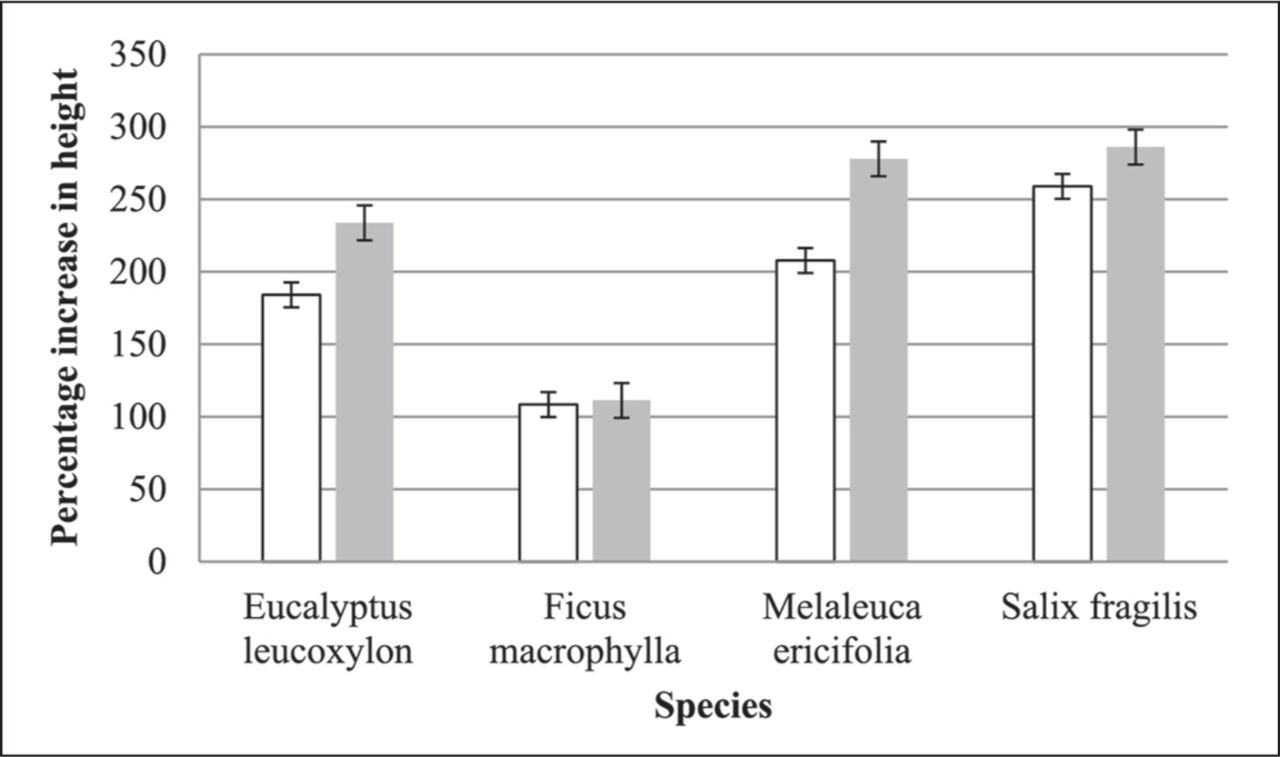
Comparing the percentage increases in height for each species irrigated by water and stormwater showed that plants irrigated by stormwater were taller than those irrigated by water, significantly so for three species (Figure 9).

Percentage increases in height for species irrigated by water (white columns) and stormwater (grey columns). Vertical bars show Least Significant Difference (P < 0.05).
Comparing the percentage height increases for different hole configurations showed that the controls were tallest. Trees irrigated with stormwater were generally taller (Figure 10).

Percentage increase in height against hole configuration for trees irrigated by water and stormwater. Vertical bars show Least Significant Difference (P < 0.05).
DISCUSSION
The experiments revealed that tree roots penetrated the narrowest cracks and smallest holes. A small number of plants penetrated 0.04 mm cracks, but subsequent root growth was poor. Both experiments confirmed differences between species in their capacities to penetrate cracks and holes and proliferate. The experiments showed that the wider the crack or the greater the number of holes, the more likely the occurrence of penetration. The greater number of holes provided more opportunity for root penetration. There was no root penetration of properly welded PVC pipe-caps in these experiments.
These experiments showed that roots penetrated through holes in polycarbonate plates as small as 0.5 mm diameter, which is consistent with Jacobs (1955), who found that Eucalyptus bicostata roots grew through 0.2 mm copper wire gauze, and that the roots which grew through the gauze were very small. Roots also grew through 1.25 mm holes drilled into the sides of pots into good soil outside the pots. Absorbing root tips measure 0.2 to 1 mm in size (Perry 1982), so the penetration of 0.04-mm cracks was unexpected and warrants further investigation. However, when a root is compressed from the sides, it can produce a greater force in elongation if it has a thicker diameter, and it has been noted that roots can narrow to pass through cracks and holes smaller than their usual diameter and then thicken once they have passed through the opening (Coder 1998; Stal and Rolf 1998).
Species such as M. ericifolia, A. littoralis, and S. fragilis, which are known for their tolerance of waterlogged soils, had strong root growth after penetrating holes greater than 0.5 mm and wider cracks. P. calleryana and C. salignus had strong root growth after penetrating the wider cracks, and C. salignus was able to penetrate the finest of cracks. F. macrophylla had strong root growth after penetrating holes of 0.5 mm diameter or larger. L. confertus and E. leucoxylon had only moderate root growth after penetrating larger cracks and holes, and while E. leucoxylon did penetrate the narrowest crack, the roots showed little subsequent growth. If the tap root of E. leucoxylon did penetrate holes and cracks, which was rare, root growth was strong. The roots of A. palmatum were able to penetrate wider cracks but made little growth.
Species from genera such as Melaleuca, Eucalyptus, and Salix are known to cause damage to sewerage and stormwater pipes (Pohls 2001), and Ficus species often damage urban infrastructure, and so their roots were expected to penetrate and proliferate through cracks and holes. There were similar expectations for species that tolerate waterlogged soils, but the experiments showed that all species had the capacity to penetrate cracked pipes. However, the poor root growth and proliferation for those plants that penetrated the narrowest cracks suggests that there may be a limit to root growth through such a narrow constriction.
While these experiments were done on seedlings, the meristematic cell division and differentiation responsible for apical shoot and root tip growth have been reported to be similar in studies on mature and juvenile plant tissue development (Sawchuk and Scarpella 2013). Because responses to phytohormones occur at a cellular level, consistent tissue responses are to be expected (Nakamura et al. 2012; Koepke and Dhingra 2013), which would suggest that the action of the roots tips of mature trees will be the same as those reported for seedlings.
Experiment 1
Many roots penetrated the wider cracks, but only five penetrated the 0.04 mm cracks. Very fine roots developed in pipes with the narrowest cracks, and longer and heavier roots grew in the pipes with medium and large crack widths. Fine cracks seemed to limit subsequent root growth, even if the crack was penetrated. There were significant differences between species in their capacity to penetrate cracks. Only three species penetrated the narrowest cracks: C. salignus and single instances for E. leucoxylon and L. confertus. C. salignus (68%), A. littoralis (60%), L. confertus (58%), E. leucoxylon (44%), and P. calleryana (43%) had higher rates of penetration compared to A. palmatum (18%).
Differences in root growth and architecture explain some of these differences in crack penetration. From observations during harvesting, C. salignus, A. littoralis, and L. confertus had many fibrous roots distributed through the pots and a high chance of encountering cracks in pipes. E. leucoxylon seedlings had long, woody tap roots which wound around the bottom of the pot with few laterals and so had a lower chance of encountering cracks. P. calleryana had several widely spaced woody roots growing from the stem and down the sides of the pots, but fewer roots in the middle of the container than species with fibrous roots, and again less chance of encountering cracks. A. palmatum had less numerous and finer roots than other species, which resulted in lower rates of penetration. Species root architecture may be exploited where pipe penetration is a problem.
Although there were four different contents of the PVC pipes, the only significant difference (P < 0.05) was in the greater root penetration and growth for pipes filled with soil compared to those filled with stormwater soil. Tree roots are opportunistic, growing where their requirements are provided (Perry 1982), and it is likely that they could proliferate in soil more easily than stormwater, as soil contains higher levels of oxygen.
Experiment 2
The controls had the greatest root mass in the lower parts of the pots, as there were no plates to restrict growth. There was no significant difference in the percentage of root penetration for different sized holes, and all four species could penetrate holes 0.5 mm or greater in diameter. It was surprising that roots of F. macrophylla could penetrate 0.5 mm holes, as their fine root diameter was larger than 0.5 mm, but their root tips passed through the holes and then thickened again.
Significant differences between species were expected, as there are clear differences in root structure and architecture. S. fragilis had the heaviest mean mass of roots in the lower parts of the pots, perhaps because their roots were already becoming lignified. F. macrophylla had heavy, rubbery roots, while M. ericifolia had fine roots. The roots of E. leucoxylon were very fine, apart from the tap root, none of which grew through holes in this experiment.
Generally, plants irrigated with stormwater were heavier and taller than those irrigated with water, and towards the end of this experiment some of the water-irrigated plants were beginning to look less healthy than stormwater-irrigated plants. The stormwater, containing 6 mg/L of nitrogen and 0.6 mg/L of phosphorous, was providing a source of nutrients (Denman 2009; Denman et al. 2011). There was an interaction between hole configuration and species for percentage increase in height, with S. fragilis and M. ericifolia growing the tallest for controls and plates with mixed hole sizes and 0.5 mm diameter holes, but these are the fastest growing species.
CONCLUSION
These experiments suggest that both pipe condition and species root architecture impact the incidence of PVC pipe penetration by tree roots. The wider the crack, or the more numerous the holes, the more likely they will be penetrated. However, root architecture also has a role to play, with species that have many fine roots being more likely to access holes and cracks than species with large, single tap or vertical roots. The use of properly installed PVC piping is likely to result in fewer pipe penetrations in future, but when planting street trees over older and deteriorating pipes, it would be wise to consider root architecture when choosing appropriate species for planting.
ACKNOWLEDGEMENTS
Graham Hepworth of the Statistical Consulting Centre, University of Melbourne, assisted with statistical analyses. Lophostemon confertus plants were donated by Mancefield Nursery. We thank the following colleagues from University of Melbourne Burnley Campus: Alicia Fitzgerald, Ross Payne, Jamie Pearson, Alex Campbell, Vince Symons, Nick Osborne, Clare Scott, Andrew Smith, and Steve Elefteriadis for assistance. Susan Bendel thanks Daniel and Jesse Bendel, Liz Dekavalas, Philip Dekavalas, John Dekavalas, Laurie Krauss, Kim Rea, and Gillian Lubansky for assistance in undertaking the experiments and their analysis.
- © 2019, International Society of Arboriculture. All rights reserved.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.