
Abstract
Trees growing in the urban environment are often subject to a variety of edaphic stressors that can lead to premature decline. As a result, soil restoration methods are required. The scoop and dump (S&D) technique of soil remediation is the practice of incorporating large volumes of organic matter (33% v/v) into the soil profile. A controlled greenhouse experiment was conducted to determine if S&D soils affected growth of Weeping Fig (Ficus benjamina ‘Evergreen’) trees during establishment (fourteen weeks) and gas exchange during a twenty-six-day water deficit period. S&D soils displayed decreased mean bulk density, increased accumulative gravimetric water holding capacity, and increased mean organic matter compared to unamended soils. In the remediated soils, Ficus benjamina ‘Evergreen’ showed a significant increase in the growth of roots, shoots, and leaf area. The increased leaf area of trees in the S&D soils caused greater transpiration on a wholeplant basis. As a result, the increased water-holding capacity in S&D soils were utilized at a statistically equal rate as the trees in unamended soils. Trees growing in both soil treatments displayed statistically equal reductions in gas exchange during the water deficit period. This study finds that urban soils treated with the S&D technique significantly reduces root-limiting compaction and increases tree biomass growth.
INTRODUCTION
Soil compaction resulting in increased bulk densities is commonly found in the urban landscape (Randrup et al. 1997; Scharenbroch et al. 2005; Gregory et al. 2006; Yang and Zhang 2011). High-bulk density soils have been shown to have adverse effects on root development, which can lead to reduced performance or premature decline of plants growing in them (Duffy and McClurkin 1974; Daddow and Warrington 1983; Jim 1993; Day et al. 2010; Alameda and Villar 2012).
The combined effects of compaction and water deficits have been shown to reduce respiration rates and root hydraulic conductivity for trees growing in urbanized landfill soils (Liang et al. 1999). Conversely, the low-bulk densities of soils (1.05 to 1.45 Mg m−3) in Singapore’s urban forests were shown as a non-limiting factor for urban tree growth (Ghosh et al. 2016).
Adequate soil volume and available water holding capacity (AWHC) are two important factors that can determine the maximum growth and longevity of trees planted in the urban landscape (Lindsey and Bassuk 1991; Lindsey 1992). City planners and landscape architects use soil volume calculations to determine the amount of soil needed for trees to reach desired size and crown projection (DeGaetano and Hudson 2000; Trowbridge and Bassuk 2004; Haege and Leake 2014). In the Trowbridge and Bassuk model, the assumption is that increasing AWHC in soils can decrease total soil volume needed to reach desired canopy density or allow for an increase in the number of trees planted in a given area.
In an effort to remediate the impact of compaction and improve the quality of urban soils, the Scoop & Dump (S&D) technique of soil remediation was developed. The S&D technique is the practice of incorporating large volumes of organic matter (approximately 33% v/v) deep into the soil profile using a backhoe, mini-excavator, or shovel (Sax et al. 2017). The practice combines the mechanical fracturing of the soil with the incorporation of organic matter. Incorporating compost into soils to improve quality has been extensively examined. Few studies, however, have focused on their effects in the urban environment (Cogger 2005). A similar technique to the S&D method, known as soil profile rebuilding, has been shown to improve soil bulk density and transplant success of trees in the mid-Atlantic U.S.A. Soil profile rebuilding shows improvements in soil quality in as little as eight months, and quality remained elevated for as long as five years (Layman et al. 2016). In other studies, the addition of compost to urban soils has been shown to have a species-specific positive effect on basal-area and stem-volume growth. The positive growth response associated with compost additions has been attributed to increased water holding capacity and microbial biomass in compost-amended soils (Oldfield et al. 2015).
Previous research on the S&D method has shown improvements in remediated soils by decreasing bulk density, increasing organic matter, and increasing AWHC within a year of compost incorporation (Sax et al. 2017). Based on these results, a study was designed to observe how trees would respond to growing in S&D remediated soils. The first objective of this study was to determine how biomass production (root, shoot, and leaf-area growth) would respond to the S&D soil environment compared to unamended controls. The second objective was to determine if trees growing in S&D and unamended soils would express differences in leaf water potential and gas exchange during a period of soil-water deficit.
MATERIALS AND METHODS
The Scoop & Dump (S&D) remediation process was conducted at the time of site establishment for landscapes adjacent to buildings on the Cornell campus. Landscapes include sites next to the Computing and Communications Center (CCC) in 2012, Mann Library in 2010, and Plant Science building in 2007. For the S&D process, Cornell compost was used and made of greenhouse medium, plant residue, manure, food waste, and animal bedding. Compost was spread across each site to a depth of 15 cm (5.9 in). A bobcat E35 Compact Excavator with a 45 cm (17.7 in) bucket was used to systematically scoop into the compost/soil layer to a depth of 38 to 45 cm (14.9 to 17.7 in) followed by lifting the material 1.2 m (3.9 feet) into the air before dumping the soil/compost back onto the ground. After the Scoop & Dump process was complete, the site was smoothed with iron rakes before plants were installed. After planting, a 7.5 cm (2.9 in) mulch layer was applied to the site. Mulch was reapplied each year between April to May until canopy closure occurred (Sax et al. 2017).
Sample Collection
Soil samples were collected from October 8, 2013 through October 15, 2013 on the Cornell Campus, Ithaca, NY, U.S.A. in the northern temperate region USDA Zone 5b. S&D samples were collected on the Agriculture Quadrangle (42.448758, −76.478115) in planting beds adjacent to buildings that had received S&D remediation in the past including the Computing and Communications Center (CCC) (42.449102, −76.479024), Mann Library (42.448692, −76.476697) and Plant Science building (42.448452, −76.477404). Unamended control soils (UN) were taken from turfed landscapes near S&D sites that shared a common land-use history.
Ten cores were extracted per study site in S&D and UN soils resulting in a total of 30 S&D and 30 UN samples. For all S&D cores, the samples were collected along a transect at regular intervals in the center of the beds. The first core of each S&D site was extracted five feet from the edge of a designated planting bed. Specific sampling intervals varied depending on the total length of the planting bed and are detailed in Table 1. Unamended soil cores were extracted at equal intervals along the transect to the S&D samples, but in adjacent turfed landscapes directly next to the study sites, and are detailed in Table 1.
Sampling location of Scoop & Dump and unamended soil cores.
Soil cores were extracted using lengths of PVC pipe that were 14.4 cm (5.6 in) in diameter and 15.0 cm (5.9 in) in height with a total volume of 2443 cm3 each. When extracting soils, the turf or mulch layer was sliced/scraped away before inserting cores into the ground. To collect UN soil, the core was slowly pushed into the ground using a hydraulic plate compactor attached to a backhoe to provide an even compression surface. For S&D soils, cores were driven into the ground using a hand tamper. After cores were tamped or pushed into the ground, they were extracted with a shovel and the core bottoms were secured with a mesh screen and ring clamp to hold the soil in place and provide drainage. After the soil cores were collected, they were brought to a campus greenhouse and growth chamber for the duration of the experiment.
Tree Growth Period
The woody evergreen plant Ficus benjamina is used as a street tree in tropical and sub-tropical regions of the world, known for its fast growing, urban- and heat-tolerant characteristics (Vargas-Garzon and Molina-Prieto 2012). Ficus benjamina has been evaluated for its ecosystem services in the urban forest such as rainfall interception (Guevara-Escobar et al. 2007) and localized cooling (Jimenez 2008). Ficus benjamina ‘Evergreen’ was selected due to its common use as an urban tree and so that this study could be conducted during winter months when northern temperate tree species are dormant (Loh et al. 2003).
A 72 cell plug tray of tissue-culture propagated Ficus benjamina ‘Evergreen’ trees were received from Sunshine Horticulture LLC (Apoka, Florida, U.S.A.) on September 23, 2013 and were acclimatized in the greenhouse for 26 days. Trees were approximately 12 cm (4.7 in) tall, with three to five stems and a root volume of 5 cm3 when they were received. On October 18, 2013, Ficus benjamina ‘Evergreen’ were planted directly into the soil cores in the greenhouse using a metal stake to dig out a planting hole. Disturbed soils were retained on the top of the cores. Resident growing medium was left attached to roots of the tree plugs. After planting, soils were watered until they reached field capacity. Five cores from each of the three sites and two treatments were selected at random and thereafter used as the study plants. Study plants were randomly distributed across a greenhouse bench. The greenhouse was heated to maintain a daytime temperature of 26.6°C (80°F) and nighttime temperature of 21.1°C (70°F). Supplemental lighting (Parsource GLXI 1000W Petaluma, California, U.S.A.) was used daily from 5am to 10pm EST with a 7-hour night period from planting October 18, 2013 until January 11, 2014. On January 11, the supplemental lighting was adjusted to illuminate trees from 6 am to 12 pm. Plants were watered daily between 8:15 to 8:45 am. The study trees received an average of 72.95 μmol s−1m−2 (+/− SD 30.33) of supplemental greenhouse lighting in addition to daily solar irradiance. Monday–Friday during the plant establishment phase, 150 ppm of N fertilizer was used in applied watering, and clear water was used Saturday and Sunday. The establishment period ended on November 13, 2013 and only clear water was used after this point.
The study plants were moved to a growth chamber on January 21, 2014. Lights in the growth chamber were set to 25% corresponding with a 16-hour day providing 5.356 to 7.085 Mol Par/16 hr. On January 29, 2014, light intensity was increased to 100% to reach between 19.41 to 26.9 Mol Par/16 hr. Day temperatures were set to 26.6°C (80°F) with the photoperiod occurring from 8 pm to 12 pm. Night temperatures were set at 21.1°C (70°F). Humidity was left at ambient levels.
On January 28, the soil depth for each container was determined by measuring the difference in height between the top lip of the container and where the soil line started at four evenly spaced locations around the pot. These measurements were used to determine total soil volume and eventually bulk density. The initial growth period ended on February 1, 2014, and all study plants were irrigated until soils were saturated at field capacity. After soils had finished draining freely, each core was weighed to determine total starting weight at field capacity. Total weight included trees, soil, PVC container, mesh screen, clamp ring, plastic bag, rubber band, and twist tie. Plastic bags were tied around the PVC containers and the top of the soil profile to reduce water loss by soil evaporation. Volumetric water content was determined using a four prong Theta Probe (ThetaProbe ML2, Delta-T Devices, London, England). All PVC cores had two sets of three holes that were drilled into the side of the containers at 4 cm (1.5 in) and 8.5 cm (3.3 in) from the top, allowing access for the Theta Probe at depth. Soil cores were weighed every five days to determine water loss between days and accumulative water. Accumulative water loss was reported in kgs on a volumetric basis due to the fact that soil and container volume were known.
Water Deficit Dry Down Period
On February 2, 2014, the water deficit dry down experiment was initiated (Day 0). The first round of sampling occurred one day later on February 3, 2014. Sampling was repeated every five days and a total number of six times with the last measurement ending on Day 26. Measurements included gravimetric and volumetric water content of containers, leaf water potential (kPa), transpiration (mmol H20 m−2S−1), vapor pressure deficit (kPa), photosynthetic rate (μmol CO2 m−2S−1), and stomatal conductance (mol H20 m−2S−1). A LI-COR LI-6400 XT portable gas exchange system (Li-Cor Inc, Lincoln, Nebraska, U.S.A.) was used and was set to provide 450 PAR light, 400 ppm CO2, 25°C (77°F) temperature and ambient humidity. Predawn leaf water potential was measured using a plant water status console pressure chamber (Soil Moisture Equipment Corporation, Santa Barbara, California, U.S.A.) (Scholander et al. 1965). For water potential testing, fully expanded leaves were removed from individual shoots with a razor blade and immediately placed into the pressure chamber. At the site of excision, naturally occurring latex was wiped away with cotton swab, so that leaf water could be observed unobstructed. A modified dissecting microscope was used to observe leaf water at the site of excision on the petiole.
The final round of sampling occurred on Day 26, and the experiment concluded. Shoots were cut at the root flare and individual leaves removed. Fresh leaves and shoots were weighed. Fresh leaves were passed through a leaf area meter (Licor LI-3100C Lincoln, Nebraska, U.S.A.) and after measurement were placed in a paper bag for drying.
Final Sampling and Experiment Conclusion
Soils were collected from the sample cores and placed in airtight plastic bags. These bags were taken to the Cornell Soil Health Lab and assessed for water holding capacity, texture, and organic matter. Soils were stored at 4°C (39.2°F) before assessment. All soils were oven-dried at 105°C (221°F) for 24 hours and then were passed through an 8 mm sieve to remove coarse organic matter. A sample splitter was used to randomize and homogenize soil samples before being tested. Texture was determined using a rapid suspension method (Moebius-Clune et al. 2016). Organic matter was assessed by loss on ignition method (LOI). Percent loss on ignition was converted to percentage organic matter and multiplied by a correction factor. The equation utilized was: % Organic Matter = (% Loss on ignition × 0.7) − 0.23 (Burt 2014; Moebius-Clune et al. 2016).
To determine water release characteristics, samples were placed in pressure plate cell rings 7 cm (2.7 in) in diameter in a Pressure Plate Extractor (Soil Moisture Equipment Corporation, Santa Barbara, California, U.S.A.). Gravimetric water content was determined at five negative pressures (−10 kPa, −30 kPa, −100 kPa, −300 kPa, −1500 kPa) after samples had reached equilibrium. Water potentials at each step of the water release curve were converted from gravimetric to volumetric water content by multiplying the value by bulk density. Available water holding capacity was determined by calculating the difference between water held at −10 kpa and −1500 kpa on both a gravimetric and volumetric basis.
Soil weight was calculated by subtracting container weight, screen weight, ring weight, fresh roots weight, fresh shoots weight, and fresh leaves weight from the total weight. Bulk density was calculated by soil dry weight and container volume.
Root systems were washed to remove excess soil, air dried for one hour, inspected to determine if water droplets were present, and then weighed to calculate fresh weight. Soil containers, metal screens, and clamp rings were weighed individually. Roots, shoots, and leaves were placed in paper bags in an oven at 70°C (158°F) for 24 hours and then measured to obtain dry weights.
Statistical Analysis
Statistical analysis was conducted using JMP Pro v.12. For normally distributed data, ANOVA tests with an α level of 0.05 were used to compare differences of mean values by treatment for bulk density, plant growth (root dry weight, shoot dry weight, total leaf area), water release characteristics, and AWHC data. A natural log transformation normalized organic matter data before ANOVA testing. Mixed random effect models were used for repeat measurements of accumulative water loss, leaf water potential, and gas exchange measurements (stomatal conductance, photosynthetic rate, vapor pressure deficit, and transpiration) over the course of the 27-day dry down period and was followed by a post-hoc comparison to determine effect of soil treatment by day. A Bonferroni correction was used to account for the effect of the mixed model in reporting p-value significance. Stomatal conductance, transpiration, and leaf water potential data were normalized using a natural log transformation. Photosynthetic rate and vapor pressure deficit data were normally distributed without transformations. Standard error was used for normally distributed data while 95% confidence intervals were reported for data adjusted by the Bonferroni correction.
RESULTS
Across the three study sites, soil texture was determined to be a loam. In S&D soils, organic matter (OM) ranged from 3.61% to 24.48% with a median value of 8.76%. In UN control soils, OM ranged from 0.85% to 3.72% with a median value of 2.51%. OM was significantly higher in S&D soils compared to UN soils (S&D = 9.49%, CI [7.37,12.22], UN = 2.32%, CI [1.80, 2.99], p ≤ 0.0001) OM. Bulk density data was significantly different between S&D and UN soils (S&D = 0.931 g/cm3, UN = 1.597 g/cm3, SE = 0.05, p ≤ 0.0001) as seen in Table 2. A linear regression analysis (R2 = 0.82, P ≤ 0.0001) showed a relationship of decreased bulk density when organic matter increased (Figure 1).
Bulk density and mean plant growth response of Ficus benajamina ‘Evergreen’ grown in Scoop & Dump and unamended soils (n = 30).

Exploratory linear regression analysis of relationship between organic matter and bulk density (g/cm3) in scoop & dump and unamended soils (n = 30) (R2 = 0.82, P ≤ 0.0001).
Plants growing in S&D soils showed greater total dry weight of roots (S&D = 6.44 g, UN = 5.21 g, SE = 0.32, p = 0.012), dry weight of shoots (S&D = 10.17 g, UN = 7.03 g, SE = 0.63, p = 0.0015), and leaf area (S&D = 933.18 cm2, UN = 579.89 cm2, SE = 59.14, p = 0.0002) (Table 2). Correlations were observed with all three growth parameters increasing as bulk densities decreased in S&D and UN soils (dry weight shoots: R2 = 0.52, p ≤ 0.0001; dry weight roots: R2 = 0.29, p = 0.021; total leaf area: R2 = 0.53, p ≤ 0.0001) (Figure 2). Positive correlations were observed, with all three growth parameters increasing as organic matter increased in both S&D and UN soils (dry weight shoots: R2 = 0.49, p ≤ 0.0001; dry weight roots: R2 = 0.32, p ≤ 0.0001; total leaf area: R2 = 0.45, p ≤ 0.0001) (Figure 3).

Linear regression of plant growth indicators (dry weight shoots [R2 = 0.52, p ≤ 0.0001]; dry weight roots [R2 = 0.29, p = 0.021]; total leaf area [R2 = 0.53, p ≤ 0.0001]) by bulk density and soil treatment (scoop & dump and unamended)(n = 30). Root limiting bulk density represents the lowest threshold of root limiting compaction at 1.40 g/cm3.

Linear regression of plant growth indicators (dry weight shoots [R2 = 0.49, p ≤ 0.0001); dry weight roots [R2 = 0.32, p ≤ 0.0001]; leaf area [R2 = 0.45, p ≤ 0.0001]) by percent soil organic matter in scoop & dump and unamended soils (n = 30).
Accumulative water loss over time was significantly different by treatment over the course of the experiment, with S&D soils losing more water compared to UN soils (S&D = 966.63 g, CI [1055.95, 884.86], UN = 653.08 g, CI [713.43, 597.84], p ≤ 0.0001) (Figure 4, Day 26). AWHC and water held at each step of the water release curve showed varying results depending on if assessment was conducted using volumetric or gravimetric measurements. Gravimetric water content was significantly higher in S&D soils for all steps on the water release curve and AWHC compared to UN soils (Table 3). Volumetric water content was not significantly different between treatments for AWHC or any of the steps on the water release curve (Table 3).
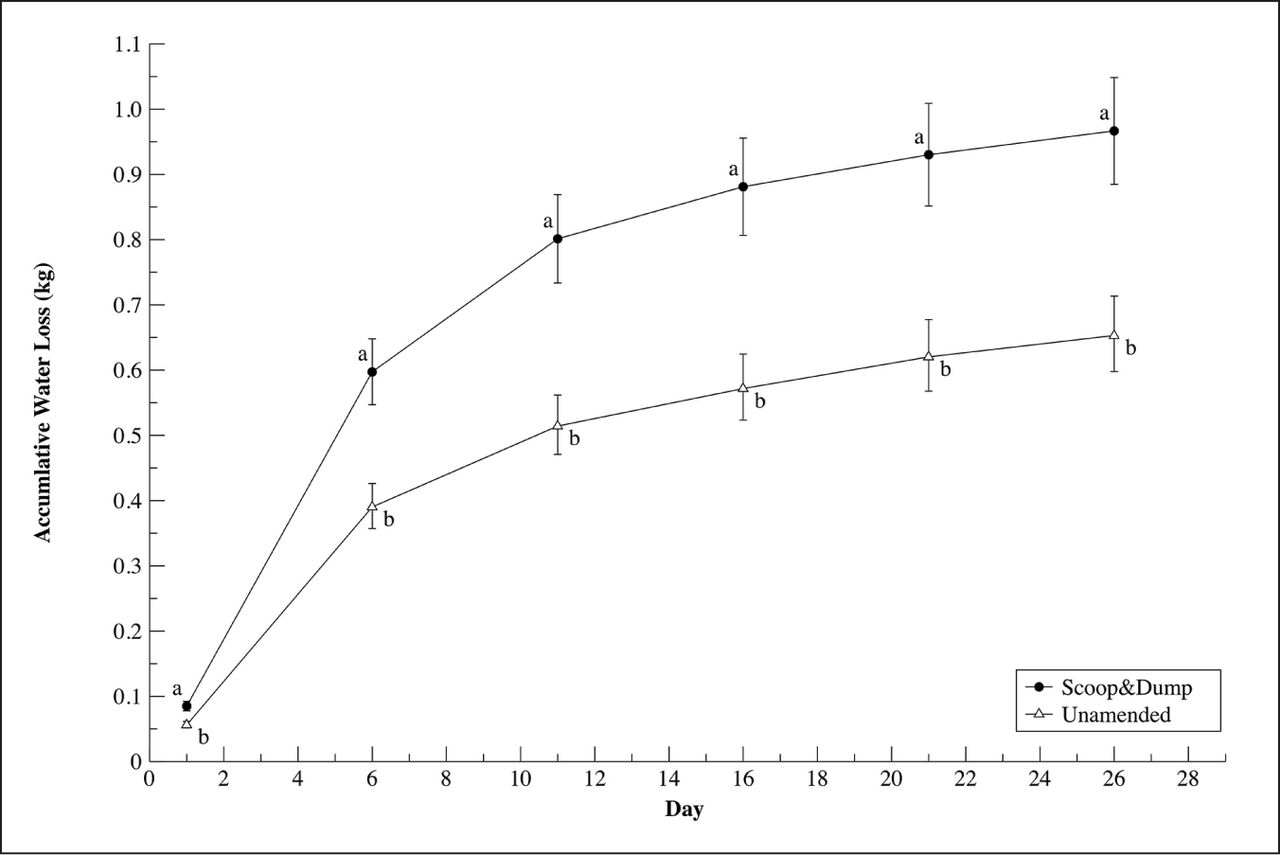
Mean values and 95% confidence intervals of accumulative volumetric water loss for scoop and dump and unamended soils over the course of a 26-day water deficit period. Containers were weighed at five-day intervals to determine gravimetric water loss between days. Containers had known volumes of soil therefore water loss is considered on a volumetric basis in kilograms.
Gravimetric and volumetric water release characteristics (θ) of Scoop & Dump and unamended soils assessed by pressure plate extraction (kPa) (n = 150).
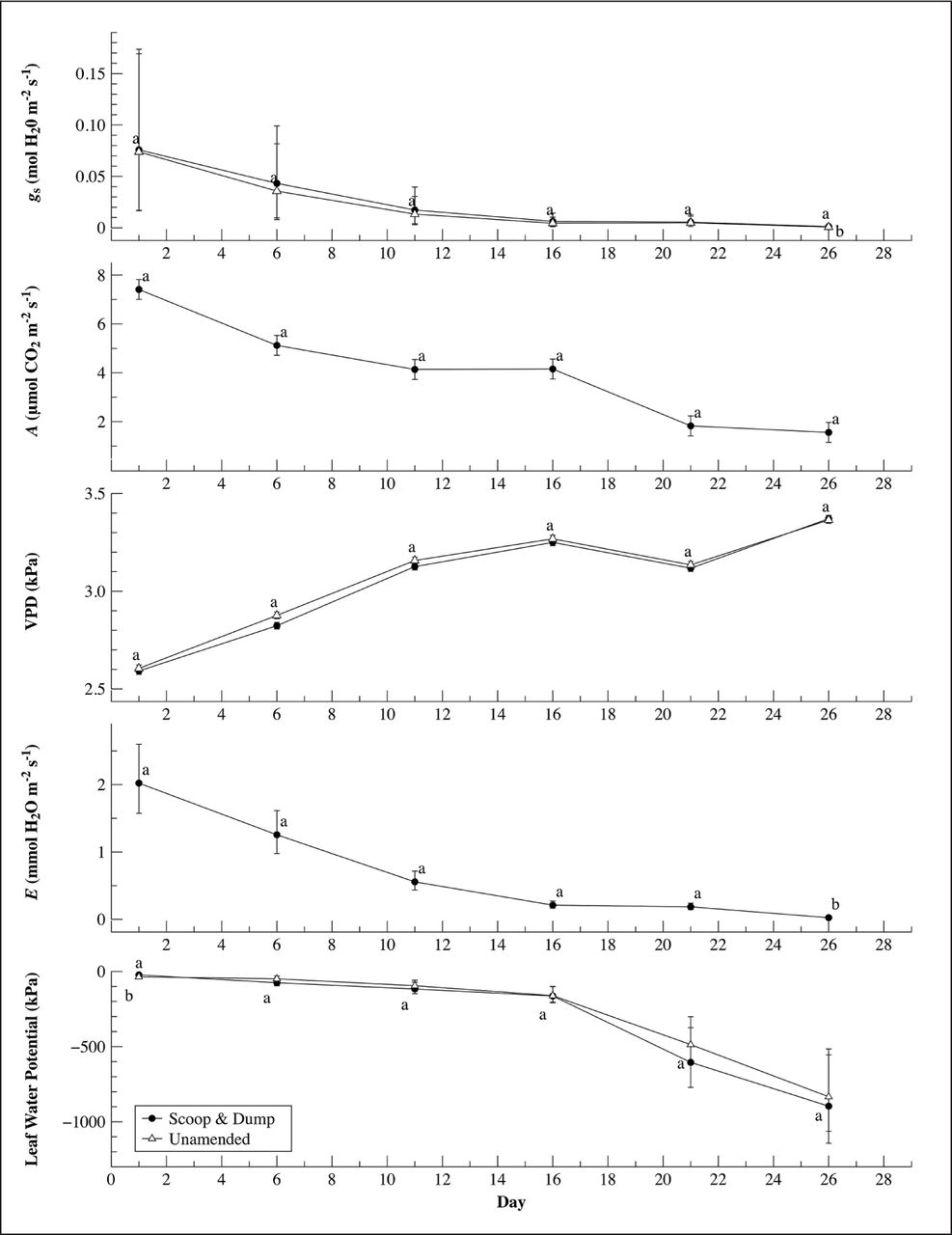
For leaf water potential, photosynthetic rate, and vapor pressure deficit measurements, no significant difference was detected between treatments over the course of the dry down period. For gas exchange measures, stomatal conductance and transpiration both exhibited statistically equal values for both soil treatments between Days 1 to 21 and unequal values on Day 26 of the dry down period (Figure 5).

Mean values and 95% confidence intervals for leaf water potential and gas exchange measurements; stomatal conductance (gs) photosynthetic rate (A), vapor pressure deficit (VPD), transpiration rate (E) of trees growing in scoop & dump and unamended soils over the course of the 26-day water deficit period.
DISCUSSION
Bulk Density, Organic Matter, and Tree Growth
This study quantified the effect of soil remediation on tree growth performance and gas exchange during simulated drought conditions. Ficus benjamina ‘Evergreen’ growing in S&D soil treatment showed a significant increase in the growth of roots, shoots, and leaf area over the course of the experiment (Table 2). The increases in plant growth correlated with decreases in bulk density (Figure 2). This result agrees with the models that Daddow and Warrington presented (1983). Through a literature review, Daddow and Warrington concluded that increased bulk densities cause a decrease in root growth and penetration. Root-limiting bulk density is texture dependent with densities of 1.45 to 1.65 g/cm3 representing the root limiting threshold for loam soils (Daddow and Warrington 1983). Duffy and McClurkin found a strong relationship between lower bulk density and planting success of loblolly pine (Pinus taeda) with 100% survival in soils with bulk densities below 1.34 g/cm3 and total failure of plantings in soils with bulk densities greater than 1.65 g/cm3 (Duffy and McClurkin 1974). Soil compaction has been shown to reduce root elongation and rooting depth in landfill soils when bulk densities were increased from 1.12 to 1.62 g/cm3, although the effects were speciesspecific (Liang et al. 1999). This result suggests that different types of trees will have a species-specific response when growing in compacted soils, and the evaluation of individual taxa is required to understand threshold levels where bulk density becomes root limiting. While a species-specific effect was detected in the Liang et al. study, root penetration for both species of trees was significantly deeper in non-compacted soils. While different species will have varying responses to growing in compacted soils, evidence from multiple studies, including this one, suggests that reducing soil-bulk densities improves overall plant performance (Figure 2) (Duffy and McClurkin 1974; Daddow and Warrington 1983; Liang et al. 1999; Layman et al. 2016). In soils remediated with a similar technique as S&D, known as soil profile rebuilding, bulk density was lower compared to the controls eight months after treatment and remained low after five years. Trees growing in profile-rebuilt soils showed increased trunk size and increased whole-tree biomass as compared to compacted control soils (Layman et al. 2016). Trees grown in S&D soils showed an increase in the growth of shoots, roots, and leaf area, demonstrating that decreasing bulk density is associated with increased plant growth in remediated soils (Figure 2). Likewise, a linear regression analysis of organic matter and plant growth showed complementary relationships with increasing root, shoot, and leaf area as organic matter increased (Figure 3). Across both unamended and S&D soils, a corelative relationship was observed with bulk density decreasing as organic matter increased (Figure 1). This relationship indicated that increased organic matter and decreased bulk density in soils can be associated with increased plant growth. While this relationship was observed in this study, neither organic matter nor bulk density alone are likely the single causal factor leading to increased plant growth. A study that looked at the S&D process over a twelve year chronosequence revealed that the S&D treatment increased soil potentially-mineralizable nitrogen, aggregate stability, organic matter, available water holding capacity, and active carbon (Sax et al. 2017). These soil health indicators combined with decreased bulk density most likely created a soil environment that caused increased plant growth in S&D soils synergistically.
Root growth in the containers was notably different between soil treatments. In S&D soils, visual examinations revealed that roots actively explored the full volume of soil in the container, growing through aggregates and pore spaces. In contrast, roots in unamended soils were primarily found directly where the original root plug was planted. When roots did grow and explore outside this region, they grew across the top of the soil and then escaped down in the space between the soil and PVC container wall. This pattern of roots taking the path of least resistance in the high-bulk density soils and not growing through the medium was consistent in all unamended soils during visual inspections. This growth response is similar to that of a study where roots were observed escaping into available pore spaces in high-density soils (Stirzaker et al. 1996). Although significant differences in mean dry root weight were observed between treatments, the author speculates that the root weight in unamended soils is greater than it would be in the landscape due to a root’s tendency to grow down the inside of the edge of the PVC containers. This artificial condition created by the PVC containers would not occur in a field setting. The use of undisturbed soil cores offers the best option for extracting and observing soils ex-situ. Despite this potential source of error, a significant difference in root weight was observable by treatment (Table 2).
Gas Exchange and Leaf Water Potential
Gas exchange measures (photosynthetic rate, stomatal conductance, vapor pressure deficit, transpiration) are commonly used to study drought response in plants because these related physiological processes are sensitive to changes over a wide range of plant water potentials (Ditmarová et al. 2009). In a water deficit study comparing physiological responses in Picea abies to three different soil water treatments, stomatal conductance, photosynthetic carbon assimilation (net CO2 exchange rate), and leaf water potential declined as the deficit period increased (Ditmarová et al. 2009). Liang et al. has shown that the combined effects of soil compaction and water defecit reduced respirations rates and root hydraulic conductivity for trees grown in urbanized landfill soils (1999). For Ficus benjamina ‘Evergreen,’ transpiration and stomatal conductance declined rapidly from Day 1 through 16 and then slowed from Days 16 through26 for the two soil treatments (Figure 5). This decline was similar to the findings of Ditmarova et al. who found a strong relationship between stomatal conductance and transpiration rates (2009). The rapid reduction in transpiration and stomatal conductance was in contrast to photosynthetic rates that had a slower decline over the dry down period, not sharply dropping at any one point in time (Figure 5).
Leaf water potential showed a slow initial decline during the dry down period and then severely dropped off after day 16 (Figure 5). Accumulative water loss showed a similar pattern of decline with the majority of the soil water being lost between days 0 and 16 (Figure 4). This pattern of accumulative water loss in the soils helped to explain the observed decline in leaf water potential.
Observing the compiled gas exchange measurements in Figure 5 shows that even small reductions in leaf water potential can result in rapid decreases in transpiration and stomatal conductance. After leaf water potential severely decreased on Day 16, transpiration and stomatal conductance fell to very low rates. Photosynthetic rates showed a large decrease in response to relatively small changes in leaf water potential. This decrease indicated that a direct relationship between leaf water potential and gas exchange should not be implicitly assumed and that in order to understand the physiological plant response to water deficit, both gas exchange and water potential measurements should be observed independently. This pattern of leaf water potential functioning at a different pace than stomatal conductance and photosynthetic carbon assimilation agrees with the findings of Ditmarova and Arndt (Arndt et al. 2001; Ditmaorva et al. 2009). Although stomatal conductance, photosynthetic carbon assimilation, and leaf water potential declined at different rates in this experiment, all three processes are linked and can function as indicators for changes in physiology of plants under water deficit. The pattern of rapid decline in stomatal conductance while maintaining predawn leaf water could illustrate the strategy of Ficus benjamina ‘Evergreen’ for coping with drought by maintaining leaf water potential by closing stomates. A similar response to water deficit conditions was reported in Ziziphus rotundifolia, which reduced stomatal conductance but maintained leaf water potential while enhancing carbon accumulation (Arndt et al. 2001).
Predawn leaf water potential (PLWP) is commonly used as in indicator for plant water status because, prior to sunrise, water status will equilibrate between soils and the plants growing in them. PLWP is considered to be a good indicator in homogenous soils that do not differ greatly across geographical space and do not contain pockets of soil with high AWHC.
PLWP and stem water potential have been shown to have greater sensitivity to changes in plant water status than midday leaf water potential (Remorini and Massai 2003). In irrigation experiments comparing different methods of measuring plant-water status, sap flow has been shown to be the strongest indicator of water potential (Remorini and Massai 2003). Sensitive measures such as cell turgor pressure (Jones 2007) have been argued as a more reliable indicator of water stress in plants. Although this may be true, the techniques required to assess cell turgor pressure are not as easily applied to field settings and typically require lab preparation (Calbo et al. 2010). Despite the limitations of predawn leaf water potential, this technique remains a relatively easy measurement to make in the field. The validity of this method may be increased if assessed jointly with other physiological indicators such as gas exchange. For example, PLWP has been shown to have a positive relationship with transpiration and can demonstrate meaningful differences in plant water status during moisture deficit conditions (Améglio et al. 1999). Figure 5 demonstrates how overlaying PLWP with gas exchange measurments can increase understading of how Ficus benjamina ‘Evergreen’ responds to water deficit conditions.
Gas exchange is highly sensitive and linked with both vapor pressure deficit (VPD) and light availability (Ditmarová et al. 2009). This relationship was demonstrated by Ficus benjamina ‘Evergreen’ showing decreased stomatal conductance when VPD increased (Figure 5) during the dry down period. This VPD response could be a compounding factor that affects transpiration and stomatal conductance when reductions in soil moisture occur.
Accumulative water loss over the course of the dry down experiment showed significant differences between treatments (Figure 4) with more water being held in S&D soils compared to unamended. The calculation of accumulative water loss was done by weighing the containers at five-day intervals, accounting for non-water weight and determining the difference in the amount of water held in the soil at the start and end of the experiment. At the end of the experiment, soil sub samples were used to asses AWHC and water release curves using a pressure plate extractor. Raw data derived from using the pressure plate extractor was reported on a gravimetric basis and is typically converted to volumetric by multiplying gravimetric water content by bulk density. Table 3 showed water release curves and AWHC reported on both a gravimetric and volumetric basis. On a gravimetric basis, significantly more water was held in S&D soils compared to unamended at all steps in the water release curve and AWHC. As a result of large differences in average bulk density in S&D (0.931 g/cm3) and unamended (1.597 g/cm3) soils, the conversion of AWHC and water release steps to a volumetric basis showed a different pattern compared to results found with gravimetric. Volumetric AWHC and water release showed no significant differences between treatments for any of the steps or AWHC (Table 3). This difference between gravimetric and volumetric measurements is illustrative to the limitations of the AWHC measurement, perhaps in this case due to large differences in organic matter or bulk density. Past studies of the S&D remediation process showed an increase in AWHC on a volumetric basis in S&D soils (Sax et al. 2017). The accumulative water loss (Figure 4) data definitively shows that more water was lost in S&D soils compared to unamended over the dry down period. Accumulative water loss data agrees with gravimetric AWHC and water release measurements (Figure 4 and Table 3) leaving the volumetric water content harder to interpret. While water release curves and AWHC represented a standard lab-based method for measurement of soil water potential, this study showed that other measures such as accumulative water loss may be more appropriate when certain soil factors (e.g., organic matter, bulk density) are outside of typical ranges.
Increases in organic matter (OM) in soils can increase AWHC (Rawls et al 2003; Alliueme et al. 2013). Large quantities of OM in the soil ranging from 1to 6% (v/v) OM can make up 5 to 25% of soil bulk volume (Hudson 1994). When incorporated into higher-density bulk soil, the relatively low-bulk density of compost has a diluting effect (Scharenbroch et al. 2005). Our study showed evidence that the compost incorporation that occurred when undertaking the S&D process may have improved both gravimetric AWHC (Table 3) and bulk density (Table 2).
Although S&D soils held more accumulative water compared to unamended soils, this increase did not significantly alter the gas exchange response of Ficus benjamina ‘Evergreen’ under soil water deficit conditions (Figure 5) as the experiment went on. Trees growing in S&D soils generated significantly greater leaf surface area and increased root and shoot growth (Table 2). This demonstrated that plants growing in S&D soils can develop greater tree biomass compared to plants in UN soils. As a result of increased total leaf area in S&D plants, we postulate that greater overall stomatal conductance occurred for S&D trees. This caused them to utilize the increased AWHC in S&D soils at a similar rate to trees growing in unamended soils with lower AWHC. Leaf water potential declined at similar rates for trees in both soil treatments, indicating that even though S&D soils held more accumulative water, this did not buffer the effect of soil water deficit for S&D trees. It is difficult to determine the exact mechanism that caused S&D trees to respond equally to water deficit conditions in terms of gas exchange as unamended trees, but the differences in leaf area and plant growth between treatments is a likely explanatory factor.
Future studies could be conducted to determine if plants of the same size and leaf area resist drought effects in soils with increased AWHC. This could be explored by conducting a similar experiment, but implementing a water deficit treatment after a significantly shorter plant establishment period or by pruning trees to have equal number of leaves and woody biomass. If the trees were a similar size and leaf area during water deficit, then the increased accumulative water held in S&D soils might lead to improved gas exchange rates for trees. Even though trees growing in S&D soils did not have an increased ability to buffer the effects of water deficit, the enhanced growth showed promising results for tree development in restored urbanized soils.
Further investigation is required to test the S&D method outside of the controlled greenhouse and growth chamber environment. Limitations to this study include the collection of soils directly from the landscape instead of creating a uniform experimental S&D soil. As a result, the soils data presented here represents field conditions at the time of sampling and should be considered as observational. Undisturbed soil cores were used in this study as a means of collecting samples in as close to field conditions as possible. While effective at maintaining soil structure, this method also created an artificial soil environment including the negative space between core walls and the soil. Likewise, due to the size and architecture of the PVC tubes, roots were confined to a maximum soil depth of 15 cm (5.9 in). As a result, growing trees in these undisturbed cores limits inference for direct comparison to trees growing under field conditions. Future studies can add to this body of knowledge by either creating S&D conditions in containers for use in a greenhouse and growth chamber study, or by creating urbanized soils and conducting S&D restoration under field conditions.
CONCLUSION
The S&D technique of soil remediation improved biomass growth on Ficus benjamina ‘Evergreen’ in remediated urban soils grown in greenhouse and growth chamber conditions. The tree growth biomass indicators were positively correlated with decreased soil bulk density and increased organic matter. Increases in soil’s accumulative water holding capacity did not significantly resist the effects of water deficit on plants growing in S&D remediated soil as indicated by gas exchange measurements and leaf water potentials. Gas exchange measures, transpiration, and stomatal conductance displayed similar water deficit response patterns for Ficus benjamina ‘Evergreen’ in both soil treatments. Increased plant size and leaf area of trees grown in S&D soils likely explain this observation with larger plants using soil water at similar rates to smaller trees in unamended soils. Results suggest that consideration of both total soil volume and soil quality (bulk density, OM, AWHC) are important to achieve desired tree growth in confined spaces. For trees growing in confined spaces, the use of low bulk density soils could result in larger trees with greater leaf surface area. This study adds to the developing body of knowledge that promotes compost as an urban soil amendment and method to remediate compaction. The S&D technique provides a tool for land managers that can be easily implemented using available technology and locally and sustainably sourced amendment material.
ACKNOWLEDGMENTS
The authors would like to acknowledge Sunshine Horticulture LLC, Apopka, FL, for their donation of Ficus benjamina ‘Evergreen’ for this experiment; the Cornell Soil Health Lab and Bob Schindelbeck for assistance in conducting soil sampling and analysis Dr. Harold van Es for reviewing the statistics and providing feedback on the data interpretation and analysis; Francoise Vermeylen of Cornell’s Statistical Consulting Unit for help with statistical analysis; and Briana Amoroso for assistance in editing.
- © 2019, International Society of Arboriculture. All rights reserved.
LITERATURE CITED









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.