
Abstract
The drilling resistance test has been widely used in tree inspections and structures since its first demonstration in Germany in 1988. A high correlation to wood density allows a correspondingly clear interpretation and reliable evaluation of the profiles in terms of wood condition. Without such a correlation, it is not clear what the profiles mean. In this study, researchers compared the profiles of a non-calibrated resistance drill with pictures of the surfaces of the drilled stem cross sections in order to find out how defects are revealed. In decayed areas, the profiles mostly dropped down and the results showed that advanced stages of decomposition and voids of significant size can be detected reliably with this device. There were statistical differences between drilling amplitude among the studied species and, in the heartwood, the values of amplitudes were superior and statistically different from those obtained in the sapwood. However, the depth of the drop, or the changes in the profiles of this device, do not allow users to differentiate between various stages of deterioration. Similarly, it is not clear if all rising profile peaks are caused by locally higher wood density, as compatibilizations zones, or by technical artefacts.
The drilling resistance measurement is a semi-destructive test in which a fast rotating drilling needle is inserted in the wood under inspection. The method has development considerably since the idea was first introduced in the 1970s (Rinn 2014); it has been widely used in tree inspections and structures since 1988 (Rinn et al. 1989). The needle must be very thin to minimize material damage; the needle, however, cannot be so thin that it could suffer from resonance or buckling problems (Tannert et al. 2013). Most machines use a needle with a 3-mm tip diameter where the cutting edges are located, and a 1.5-mm diameter shaft (Tannert et al. 2013). With the rotation, the needle cuts the wood and perforates the material as it advances (Botelho 2006). The tested drills require manual adjustment of the advance speed of the needle (cm/min), as well as its rotation speed (turns/min). For those drills, this adjustment must be done according to the hardness or the density of the wood (Nutto and Biechele 2015). Besides the velocities, the sharpening of the needle tip is also very important to reduce shaft friction, which causes interpretation problems of this kind of drill results, especially in dense woods (Nutto and Biechele 2015). While drilling, the needed energy is measured depending on the drilling depth of the needle, and registered in a percent-amplitude graph.
The different resistance drills differ strongly in many ways: in handling and practical applicability, in precision and accuracy, and most important, in how their profiles have to be interpreted (Rinn 2012). When drilling at the same spot of a tree with different kinds of resistance drills, the profiles can differ so strongly that users can come to contradictory evaluations of wood condition (Rinn 2015). Consequently, there is need for a criterion for how to evaluate the accuracy and reliability of the profiles obtained by the various kinds of resistance drills in terms of wood condition.
Considering that the equipment measures the resistance offered by the wood to drilling, it is also expected that the equipment’s results can be correlated with losses in density caused by deterioration. Several studies have focused on evaluating the correlations between the amplitude of the drilling resistance obtained by the equipment and the wood density (Botelho 2006) or its effectiveness in detecting deteriorations in wood structures (Eckstein 1994; Brashaw et al. 2011; Rinn 2012; Tannert et al. 2013; Rinn, 2016), logs (Brashaw et al. 2011), trees (Johnstone et al. 2007; Wang et al. 2008; Kubus 2009; Johnstone et al. 2010), and damaged wooden poles (Jenkins 2011; Reinprecht and Supina 2015).
Density is a wood material property that describes wood condition, and it differentiates between intact and decayed parts (Means et al. 1985). How important it is to differentiate between the various stages of decomposition becomes clear when realizing that, depending on the type of decay, a loss in density by only 10% can lead to a loss in strength of 90% or even more (Wilcox 1978). Structural stability of wood is thus highly dependent on even slight changes in wood density. Therefore, many research projects have focused on developing a method for non-destructive assessment of the density profiles of standing trees and timber (Rinn 2016). Shortly after the first international press releases described how a German physicist (F. Rinn) drilled thin needles into trees in order to inspect for defects by measuring density (Schubert 1989), many companies worked to develop similar devices. For a few electronically recording resistance drills, a high correlation (r2 > 0.9) of the profiles to wood density was found, even in standing trees (Brashaw et al. 2013); and because density is the most important material parameter for this technique, there is a need for studies to evaluate the accuracy and reliability of the profiles obtained by non-calibrated resistance drills. For resistance drills using mechanical spring-driven recording, the correlation to wood density was found to be weak (r2 ~0.5; Johnstone et al. 2011), making it correspondingly difficult to evaluate wood condition based on such profiles. Due to this weak correlation, it is impossible to identify and differentiate various stages of decomposition in these profiles (Johnstone et al. 2011). Consequently, only big defects with advanced stages of decay or voids can be identified reliably. For other resistance drills, it is not yet known how well the measured values correlate to wood density, although these drills are frequently used on the market, and have been for several years. Thus, it is not yet clear how to interpret these profiles correctly in terms of wood condition.
To fill this knowledge gap, researchers compared profiles of a non-calibrated resistance drill with pictures of the surface of the drilled stem cross-sections, with a special focus on how the different stages of decomposition are revealed.
MATERIALS AND METHODS
Sampling was composed of logs collected from six tree species: Centrolobium sp., Tabebuia ochracea, Liquidambar styraciflua, Platanus spp., Caesalpinia pluviosa, and Copaifera sp., widely spread in the urban arborization of São Paulo State, Brazil. Six discs approximately 200 mm high were removed from these logs. The discs were conditioned in a place with temperature and relative humidity (RH) controlled (temperature around 25°C and RH around 65%) until reaching equilibrium moisture. When they reached the equilibrium, species heartwood and sapwood presented moisture content around 10% and 11%, respectively. It is important to note that even in green condition, hardwoods tend to exhibit smaller differences in moisture between heartwood and sapwood than softwoods, and in some species, heartwood moisture is higher than sapwood moisture (Haygreen and Bower 1989). Eight equidistant points were marked on each disc perimeter for measurements with the drilling resistance equipment (IML PD 400, Wiesloch, Germany) (Figure 1). The measurements were performed in the perpendicular direction to the grain in the 8 marked points (measurement routes), obtaining 48 graphs of amplitude of drilling resistance. To help with the test, the discs were fixed to a concrete table with sergeants (Figure 1).

Drilling-resistance test on discs.
The species used in the research had different densities and deterioration conditions (Table 1). The model of equipment used needs the manual choice of different levels of feed speed (from 15 cm/min to 200 cm/min) and needle rotation (1,500 rpm at 5,000 rotations/min). This adjustment is usually made according to the expected but mostly unknown density of the wood. In light woods, the feeding speed must be higher to provide sufficient amplitude so that the curve variations can be seen. In denser woods, the feed speed must be reduced and the rotation increased so drilling is possible. Considering these aspects of the equipment, feeding and rotation velocities were adopted, manually, according to the expected wood density, and it was also necessary to consider the deteriorating condition of the disc to prevent the needle from breaking (Table 2). Other kinds of resistance drills automatically adapt the drilling speed to wood density while drilling. The user does not need to adjust the drilling speed before drilling with these devices.
Short description of disc deterioration.
Values of basic density (ρbas), feed velocity (VA), and rotation speed (VR) adopted for each species.
The wood condition influenced the choice of feed velocity. Although the species Liquidambar and Platanus have practically the same density, the feed velocity used in the Liquidambar was lower than in the Platanus (Table 2). The feed speed used in the Copaifera sample could also be 100 cm/min, but it was necessary to reduce it and increase the rotation so that the drill inlet would not break (Table 2).
After the drilling-resistance tests were completed, the discs were cut to the exact position of the drill paths. These sections were detailed through photographic record in order to compare the amplitude plots of drilling resistance with the different zones of the wood through which the drilling occurred. The equipment model used in this research provides two curves, one showing power needed for feedings of the needle, and the other for the rotation of the needle. However, older models provide only the feed curve. Nutto and Biechele (2015) observed that in high-density wood, the feed amplitude tends to increase with the drilling depth because of friction. This increase in amplitude may cause wrong interpretation of the results, since it can camouflage the amplitude drop that would be caused by deterioration.
The discs were macroscopically analyzed to identify the heartwood, sapwood, and deteriorated areas. Descriptions of the species were obtained in the literature (Freitas 2012; Lima et al. 2015; IPT 2016), as well as the support of an expert in visual assessment. For the species in which it was possible to distinguish the heartwood and sapwood regions, the average amplitude of drilling resistance obtained separately in the bark, heartwood, sapwood, and deterioration zones was determined.
For all discs, the drilling-resistance graphs obtained on each route were compared with the photographic images, by image overlay. In the case of discs with heart and sapwood zones, the average amplitudes of rotational drilling resistance were statistically analyzed using multivariate variance analysis. In this research, only the amplitude plots of rotational drilling resistance were used because the use of the two graphs (thrust and rotational resistance to drilling) made the image overlay too difficult.
RESULTS AND DISCUSSION
In the Centrolobium, Platanus, Caesalpinia, and Copaífera discs, it was possible to define the heartwood and sapwood zones due to their coloring distinction (Figure 2), whereas in Tabebuia and Liquidambar species, this distinction was not possible (Figure 2). The characteristics of the heartwood and sapwood differentiation as a function of color differentiation were similar to those proposed in the literature (Freitas 2012; Lima et al. 2015; IPT 2016). Not all of the discs contained voided areas and not all of the discs differed in types and levels of deterioration (Table 1).

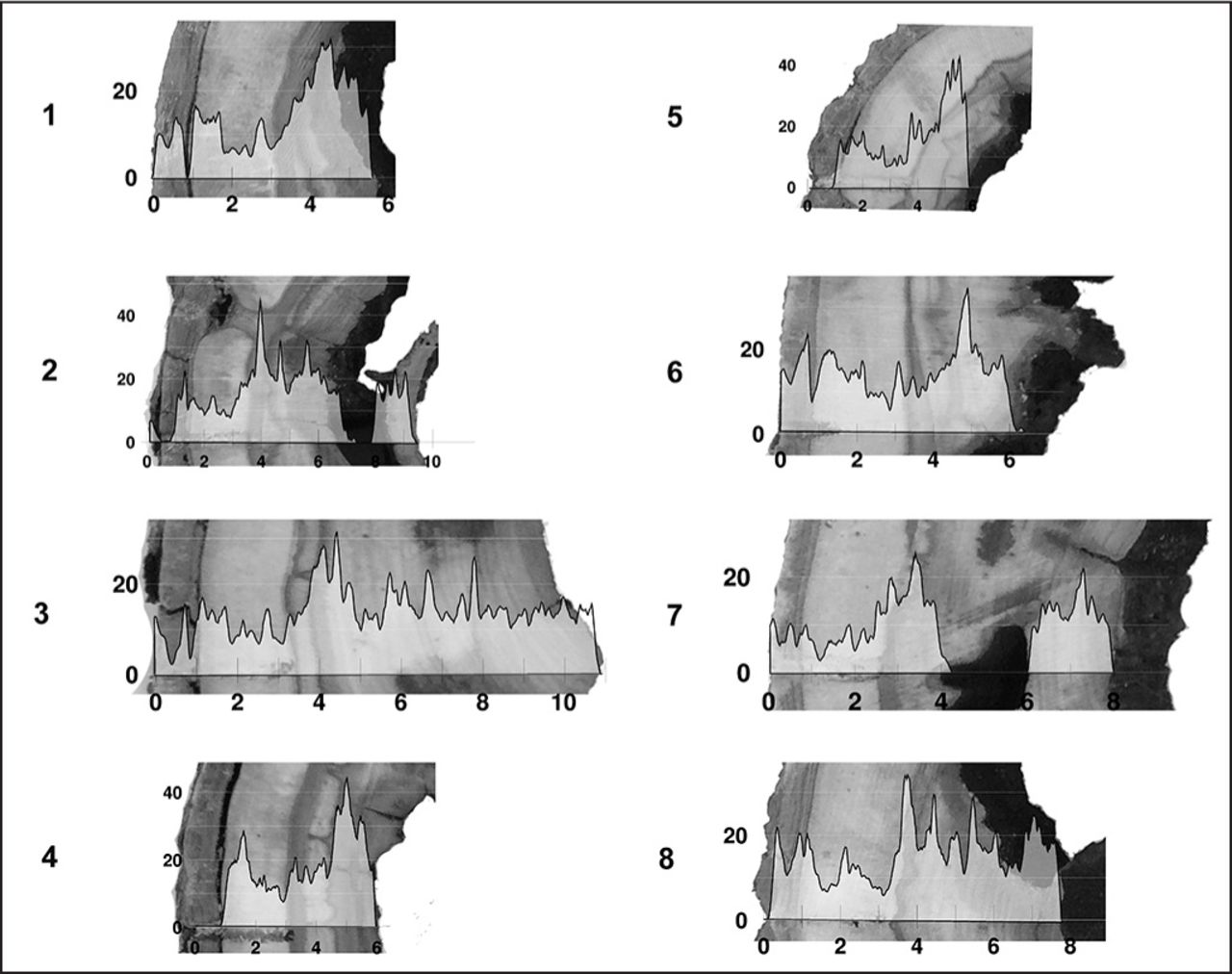
Images of the Cetrolobium sp., Platanus spp., Caesalpinia pluviosa, and Copaifera sp. discs with color distinction between heartwood and sapwood (Species Group A) and of the species Tabebuia ochracea and Liquidambar styraciflua, without distinction of color between heartwood and sapwood (Species Group B).
Considering the methodology, it was possible to obtain eight images of the amplitude graph of the drilling resistance from each disc, corresponding to the eight measurement routes, which were overlaid with the photographic images (Figure 3). The same result observed in Figure 3 was obtained for the six species studied.

Superposition of the amplitude graph of drilling resistance to the photographic image obtained from the disc, in the position of the needle passage of the resistograph. Species: Centrolobium sp.
The detailed analysis of the 48 superimposed images allowed researchers to verify the existence of a pattern of behavior of the amplitude of drilling resistance against the different zones of the wood (healthy and with deterioration) traveled by the needle during the drilling.
Considering all studied species, the maximum amplitude ranged from 0% to ≌ 40%, with the 0% sections corresponding to the voids, as expected. Considering the type of machine used (no density calibration), the amplitude range varies among species, being smaller for Centrolobium sp. and Copaifera sp. (0% to ≌ 16%) than for Caesalpinia pluviosa and Platanus (0% to 40%), which was also expected. This result confirms that this tool is adequate to detect the location and the approximate size of voids. The same result was highlighted by Kubus (2009) when analyzing the condition of a large monumental tree in Portland, Oregon, U.S. The author presents records of 10 graphs of resistance to drilling, obtained in different positions of the tree, in which there were several zones with amplitudes close to zero (probably voided zones) and, in the other regions, average amplitudes of resistance to perforation of 16%. In the bark area, the average amplitudes were 6%. Brashaw et al. (2011) obtained drilling resistance that varied from 0% to 25% in zones with different levels of wood degradation.
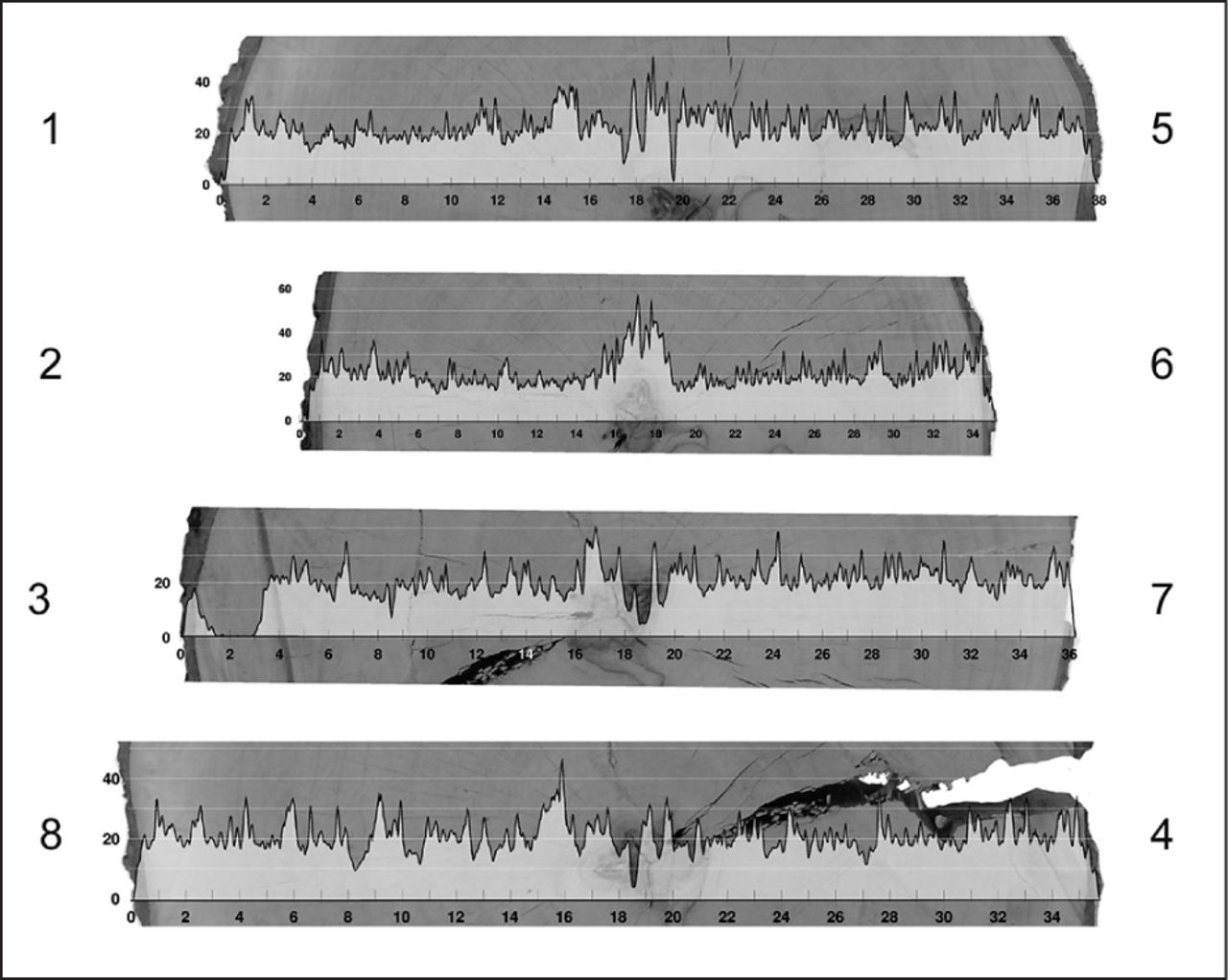
In this research, in the species for which it was possible to define the heartwood and sapwood regions, there was amplitude variation (Figure 3). In the two species for which it was not possible to visualize the distinction between heartwood and sapwood (Tabebuia ochracea and Liquidambar styraciflua), the amplitude of the drilling-resistance graph also does not show variations (Figure 4). Similar graphs, without heartwood and sapwood visual distinction, were obtained by Kubus (2009).

Superposition of the amplitude graph of drilling resistance to the photographic image obtained from the disc, in the position of the needle passage of the resistograph. Species: Liquidambar styraciflua.
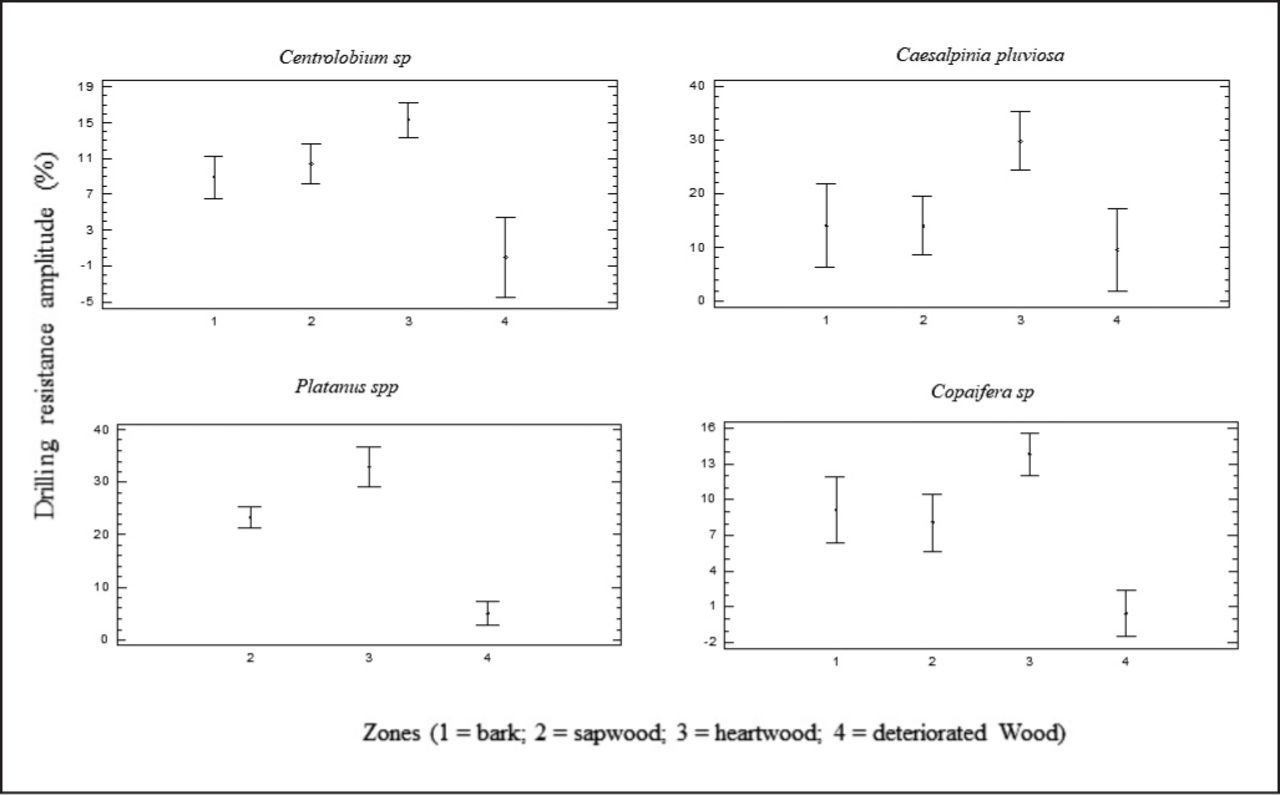
In the heartwood, the values of amplitudes were higher than those obtained in the sapwood. For the four species for which it was possible to define the heartwood and sapwood regions (Figure 2), this result was statistically demonstrated (Figure 5). The analysis of variance showed that amplitudes were always lower in deteriorated regions (with or without voids), but always higher in the heartwood regions and in sapwood and bark intermediate regions (Figure 5).

Representative graph of the average values and the variability of amplitude obtained in the different zones of the discs in different species.
Considering only the heartwood and sapwood regions without apparent deterioration, there was a differentiation between species. For sapwood, the amplitudes of drilling resistance were higher and statistically different for Platanus spp. and statistically equivalent for the other species (Table 3). For heartwood, the amplitudes of drilling resistance were also higher for Platanus spp., but statistically equivalent for Caesalpinia pluviosa, and smaller for the other two species (Table 3). This result is not expected, since Platanus spp. has the lowest density (Table 2) and the drilling resistance, in general, has a positive correlation with density (Costelo and Quarles 1999; Couto et al. 2013). However, it is noteworthy that the amplitude of drilling resistance in this device type is affected by the selected advance velocity, which for Platanus spp. was higher (Table 2), and that in order to properly interpret and compare amplitude results obtained in the resistance test, it was necessary to know, in advance, the profiles obtained in the whole condition (Martinez 2016; Matheny et al. 1999). This is different from resistance drills automatically adopting the speed to the wood, as tested by Brashaw (2013), and resulting in a high correlation to wood density (R2 > 0.9), because a high correlation to wood density is necessary to allow for reliable evaluations of the profiles concerning the wood condition.
Average drilling resistance amplitude by species and Multiple Range Test for analyzing statistical differences.
For Tabebuia ochracea, which sustained a beetle insect attack (Figure 2), the amplitude of drilling resistance did not show variations that allowed, in an inspection, researchers to visualize the wood state (Figure 6).

Superposition of the amplitude graph of drilling resistance to the photographic image obtained from the disc, in the position of the needle passage of the resistograph. Species: Tabebuia ochracea. Deterioration caused by Coleoptera insects.
In the Liquidambar styraciflua disc there was a significant crack (Figure 2) that was not identified in any drilling resistance amplitude graph, because no needle route passed through the crack site (Figure 4). This result highlights the fact that this type of inspection is punctual, and it is important to know where it should be applied. Botelho (2006) and Tannert et al. (2014) had the same conclusion.
It was possible to observe in some discs the contour of the fungi deteriorated zone, a darker-colored region. In these cases, an increase peak in the amplitude of the drilling resistance was observed (Figure 7). A similar result was obtained by Rinn (1996) and was explained by the compartmentalization process promoted in the trees’ tissue to protect them from the advance of the deteriorated zone. In this process, the tree develops a thicker cell wall tissue, in addition to obstructing cell voids, causing this tissue to act as a deterioration barrier (Shigo 1977; Shortle 1979; Fraedrich 1999). The darker color is due to the antimicrobial substances produced (Shigo 1977; Shortle 1979). The Platanus spp. disc was the only one that presented these peaks in the sapwood zone, which may have influenced the average value of amplitude of drilling resistance, making this species present the highest values of amplitude, although with lower density. Compartmentalization is an assertive hypothesis for the behavior of the amplitude resistance; however, seeing as researchers did not know the correlation of the profile with density (non-calibrated device), it is not possible to affirm that all rising profile peaks are caused by this higher wood density.

Details of amplitude peak observed in zones close to fungal decay: a) Centrolobium sp., b) Platanus spp., c) Caesalpinia pluviosa, d) Copaifera sp.
Rinn (1996) observed that differentiated patterns of behavior of the drilling resistance plot may help the inspection interpretation. The author clarifies that in fungi-deteriorated wood, the amplitudes are low and approximately homogeneous, while in termite-deteriorated areas or with cracks present, the graph is more variable and has higher amplitudes (surrounding wood in good condition) followed by amplitude values much smaller and localized. In the Platanus spp. and Caesalpinia pluviosa discs, in which there was a large fungus-deteriorated zone (Figure 2 and Table 1), the drilling-resistance graph shows a continuous segment of low amplitudes (Figure 8), according to the behavior indicated by Rinn (1996).

Superposition of the amplitude graph of drilling resistance to the photographic image obtained from the disc, in the position of the needle passage of the resistograph. Species: a) Platanus spp., b) Caesalpinia pluviosa.
CONCLUSIONS
For all studied species, the amplitude ranged from 0% to 40%, with the 0% sections corresponding to the voids, as expected, which confirms that this tool is potentially adequate for the detection of the location and the approximate size of voids. In the heartwood, the values of the amplitudes were superior and statistically different from those obtained in the sapwood, both with moisture content around 10%~11%. There were also statistical differences between species. The variance analysis showed that the amplitude in deteriorated regions (with or without voids) was always lower, except in the Coleoptera attacked wood, whose deterioration was not captured by the equipment. In some discs, amplitude-increase peaks observed on the drilling resistance graph were probably associated with compartmentalization zones promoted in the tree tissue to protect themselves from the advancement of the deteriorated zone. This behavior, also observed by other researchers, can affect the inspection interpretation by hiding deterioration, for example.
The impact of feed rate, rotational speed, and density correlation on the obtained results is critical and must be considered in further research. Curves of the same piece of wood drilled with a different feed rate and rotational speed can differ strongly, making it difficult to clearly interpret the meaning of the result. In the same way, analysis of correlation between the device profiles and real local wood density along the drilling path is necessary in order to clarify analyst interpretation and reliable evaluation of the profiles in terms of wood condition by this device.
Acknowledgments
The authors would like to thank the National Council for Scientific and Technological Development (CNPQ) for the scholarship and FAPESP (Proc. 2015/05692-3) for the research funding. They also thank the Environment Directory of UNICAMP for donating the logs used in the tests, and PD Instruments for the help and equipment used in the tests of resistance to drilling.
- © 2019, International Society of Arboriculture. All rights reserved.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.