
Abstract
Bark beetles carry a number of associated organisms that are transferred to the host tree upon attack that are thought to play a role in tree decline. To assess the pathogenicity to western white pine (WWP; Pinus monticola) of fungi carried by the mountain pine beetle (MPB; Dendroctonus ponderosae), and to evaluate the potential for systemic prophylactic treatments for reducing fungal impacts, experiments were conducted with WWP seedlings to meet three objectives: 1) evaluate pathogenicity of two MPB-associated blue-stain fungi; 2) evaluate phytotoxicity of tree injection products; 3) evaluate the anti-fungal activity of tree injection products, in vitro and in vivo, toward the associated blue-staining fungi. To evaluate pathogenicity, seedlings were inoculated with Grosmannia clavigera or Leptographium longiclavatum, common fungal associates of MPB. Seedling mortality at four months after inoculation was 50% with L. longiclavatum and 90% with G. clavigera, both significantly higher than controls and thereby demonstrating pathogenicity. Phytotoxic effects of TREE-äge®, Alamo®, and Arbotect® were evaluated by stem injection; no phytotoxic effects were observed. Anti-fungal properties of the same three products were evaluated in vitro against G. clavigera, where Alamo was most active. Co-inoculation of G. clavigera and L. longiclavatum into seedlings after a stem injection of Alamo showed significantly less mortality and lesion formation than either species alone. Results support the hypothesis that MPB blue-stain associates, particularly G. clavigera, promote death of WWP when attacked by MPB. These findings suggest that the administration of a fungicide with insecticide for tree protection against bark beetles may be advantageous.
- Bark Beetles
- Blue-Stain Fungi
- Emamectin Benzoate
- Grosmannia clavigera
- Leptographium longiclavatum
- Mountain Pine Beetle
- Pinus monticola
- Propiconazole
- Systemic Fungicide
- Systemic Insecticide
- Thiabendazole
- Tree Injection
- Western White Pine
Dendroctonus ponderosae, the mountain pine beetle (MPB), is an aggressive and destructive pest of many pine species in western North America. Lodgepole pine (Pinus contorta Douglas var. latifolia Engelmann) is a favorite host, while MPB is also a serious pest of ponderosa pine, P. ponderosa Douglas ex C. Lawson, whitebark pine, P. albicaulis Engelmann, western white pine, P. monticola Douglas ex D. Don, and limber pine, P. flexilis James (Tsuneda and Hiratsuka 1984; Yamaoka et al. 1990; Solheim and Krokene 1998; Carroll et al. 2003; Ono 2003; Lee et al. 2006a). Recently documented MPB outbreaks in western North America have resulted in millions of tree deaths (Carroll et al. 2003; Ono 2003) and reinvigorated interest in control options.
Adult MPBs mine the inner bark, moving from the attack site in an axial and planar orientation in host trees. High attack densities and the successful establishment of brood, which mine laterally, result in tree girdling and subsequent host death. Successful reproduction by MPB depends on its relationship with associated fungi (Raffa and Berryman 1983; Yamaoka et al. 1990; Yamaoka et al. 1995; Six and Paine 1998; Rice et al. 2007b). The three main blue-stain ascomycetes isolated from D. ponderosae are Grosmannia clavigera (Robinson-Jeffery and Davidson) Zipfel, de Beer and Wingfield [≡ Ophiostoma clavigerum (Robinson-Jeffery and Davidson) Harrington], Ophiostoma montium (Rumbold) von Arx, and Leptographium longiclavatum Lee, Kim and Breuil (Tsuneda and Hiratsuka 1984; Yamaoka et al. 1995; Solheim and Krokene 1998; Kim et al. 2005; Lee et al. 2005; Lee et al. 2006b). These fungi are not only vectored by MPB but are also considered to have a mutualistic relationship with this insect. It is suggested that these fungi aid the beetle in overwhelming host defenses (Raffa and Berryman 1983), providing nutrition (Six and Paine 1998; Adams and Six 2007), and perhaps a more favorable environment for beetle development (Lee et al. 2006a). For example, axenic MPBs are capable of entering and mining host trees, but offspring are apparently insufficiently nourished and attacks do not result in successful reproduction (Six and Paine 1998). In addition, fungal symbionts of MPBs have been shown to cause tree mortality by themselves (Mathre 1964; Basham 1970; Shrimpton 1973; Strobel and Sugawara 1986; Yamaoka et al. 1995). It appears that the combined action of the beetle and the fungi result in a more rapid host death (Amman et al. 1989; Yamaoka et al. 1995; Solheim and Krokene 1998; Lee et al. 2006a).
In a study using western white pine (WWP), P. monticola, trees in situ in northern California, U.S., TREE-äge® (4% [wt/wt] emamectin benzoate (EB), Syngenta Crop Protection, Greensboro, North Carolina, U.S.) injected trees had significantly reduced adult MPB gallery length compared to untreated trees (10.1 ± 2.1 and 120.6 cm ± 13.4 SE, respectively), and did not appear to be girdled by adult beetle activity (Strom et al. unpublished). However, significant tree mortality was observed in the study. To investigate potential causes of tree death, study trees were felled, wood samples collected, and fungal isolates cultured. The isolates were submitted for identification to Drs. Michael Wingfield and Wilhelm de Beer of the Forestry and Agricultural Biotechnology Institute (FABI), Pretoria, South Africa, who confirmed isolates were two species: G. clavigera and L. longiclavatum, both of which are common associates of MPB (Lee et al. 2005; Lee et al. 2006b).
In this study, the authors consider two factors that may impact tree survival: 1) injected formulation and 2) MPB fungal associates. Applied correctly, tree injections have been found to be both useful and effective against insects and pathogens. Doccola et al. (2011b) demonstrated that green ash (Fraxinus pennsylvanica Marsh.) successfully compartmentalized tree injection sites following treatment with TREE-äge for emerald ash borer (Agrilus planipennis Fairmaire). Furthermore, hemlock woolly adelgid (Adelges tsugae Annand)-infested eastern hemlock (Tsuga canadensis L.) recovered with new growth following imidacloprid tree injection (Doccola et al. 2012). Other researchers have not documented tree mortality from injections, and have generally found little or no phytotoxicity with currently available products when used according to label (Grosman et al. 2002; Smitley et al. 2010; Doccola et al. 2011a; Doccola et al. 2011b; McCullough et al. 2011).
Although Ophiostomatoid species vary in their pathogenicity to trees, a few do kill trees and this is especially true of species exotic to the host. Among these are Ophiostoma novo-ulmi (causal agent of Dutch elm disease), Ceratocystis fagacearum (Bretz) Hunt (causal agent of oak wilt), and Raffaelea lauricola (causal agent of laurel wilt). In addition, the native black stain root disease (Leptographium wageneri) can be a lethal vascular wilt pathogen of conifers in California, U.S. (Wagener and Mielke 1961). All of these species are vectored by insects that bore into trees. Furthermore, MPB-associated fungi were reported to cause injury and mortality following inoculations in lodgepole pine, jack pine, and lodgepole × jack hybrids (Strobel and Sugawara 1986; Yamaoka et al. 1990; Yamaokoa et al. 1995; Lee et al. 2006b; Rice et al. 2007a; Rice et al. 2007b; Rice and Langor 2009). It is suggested that once fungal associates colonize the sapwood, they impede water and mineral transport in these tissues (Amman 1978). However, the authors are not aware of previous research regarding the virulence of G. clavigera and L. longiclavatum in P. monticola.
In this study, researchers report on the effects of systemic injections of TREE-äge, Alamo, and Arbotect to WWP seedlings and the subsequent challenge of these treatments via inoculations with G. clavigera or L. longiclavatum. The effects of the three systemic pesticides on the in vitro growth of G. clavigera were also investigated.
METHODS
Fungal Isolation
Trees cut from a field study in the Modoc National Forest, Alturas, California, U.S., were the source of wood samples used in the fungal isolations. Sample chips were removed from wooden block samples of TREE-äge-treated and untreated trees as described in Shigo (1986) using a No. 11/4 concave wood gouge. Chips were placed into Difco™ Malt Extract Agar (MEA) and incubated in the dark at 25°C for one week. Samples were then examined for Ophiostomatoid fungi by observing characteristic conidiophores. From this isolation, two species were differentiated based on their morphology, G. clavigera by a more highly branched colony with more elongate and clavate shaped conidia, and L. longiclavatum by a less frequently branching colony with smaller and more cylindrical conidia (Six et al. 2003). Confirmatory identifications by DNA sequencing were conducted by Drs. Wingfield and de Beer. These isolates are now maintained in the culture collection (CMW) of FABI, under G. clavigera #38988 and L. longiclavatum #38989.
Fungicide Screening
Three systemic injection products including TREE-äge (4% [wt/wt] emamectin benzoate, Syngenta Crop Protection, Greensboro, North Carolina, U.S.), Alamo (14.3% [wt/wt] propiconazole, Syngenta Crop Protection, Greensboro, North Carolina, U.S.), and Arbotect (20% [wt/wt] thiabendazole, Syngenta Crop Protection, Greensboro, North Carolina, U.S.), were evaluated in vitro to determine their effect on growth of G. clavigera. To do this, serial dilutions of each treatment were established, from 10,000 to 1 ppm, along with a sterile water control. Individual filter papers, previously sterilized in an autoclave and left to dry in a sterile environment (1.5 cm, VWR, Radnor, Pennsylvania, U.S.), were dipped halfway into respective treatment solutions with sterile forceps and placed flat at the outer edge of an MEA agar plate, with the edge of the paper ~2 cm from the center of the plate (two filter papers per plate, three plates per treatment). Each plate medium was then streaked in the middle with conidia from a one-week old colony of G. clavigera. Researchers did not include L. longiclavatum in the screening because colonies did not survive cold storage. Plates were incubated at 25°C for one week, after which time measurements were made, and the experiment was concluded. The distance from the edge of each filter paper to the closest viable mycelium was measured (repression zone). If contamination obscured the view of the filter paper, no measurement was taken.
Mean repression of fungal mycelium was calculated in millimeters from six measured repression zones, two per Petri dish, for each treatment. Analysis of variance (ANOVA) for a hierarchical design with nested errors was implemented using the MIXED procedure in SAS (V. 9.3, SAS Institute Inc., Cary, North Carolina, U.S.) with treatment as a fixed effect and dish within treatment as random, followed by comparisons among treatments using Fisher’s protected LSD. A second analysis was performed on the subset of treatments obtained by eliminating the control to test for main and interaction effects of the factors fungicide and concentration. Demonstration of a highly significant fungicide by concentration interaction on repression was followed by orthogonal linear contrasts (via Estimate statements) to test for a response to increased concentration (on a log scale) of each fungicide.
SEEDLING EXPERIMENTAL OVERVIEW
Following the screening of fungicides in vitro, two experiments with seedlings were conducted. The objectives of the first experiment were to 1) test the pathogenicity of G. clavigera and L. longiclavatum in WWP seedlings, and 2) test the phytotoxicity of systemic pesticides in WWP seedlings. The objective of the second experiment was to test whether multiple inoculation points of fungi altered results compared to a single inoculation.
Experiment #1
Pathogenicity of fungal isolates
Pinus monticola seedlings were purchased from the University of Idaho, Franklin H. Pitkin Forest Nursery (Moscow, Idaho, U.S.), and potted into 15.24-cm pots using a 1:2:1 mix of compost, peat and perlite. From this group of seedlings (average basal diameter = 0.67 cm ± 0.01 SE and average height = 16.8 cm ± 0.5 SE), 50 individuals were selected and randomly assigned to 5 treatments (n = 10 replicates per treatment). The five treatments were 1) G. clavigera inoculation, 2) L. longiclavatum inoculation, 3) co-inoculation of G. clavigera and L. longiclavatum after injection of systemic fungicide Alamo, 4) sterile MEA, or 5) de-ionized water. No seedlings were inoculated with a mix of both fungi only. It is known that these fungi are commonly associated and occur in common in nature. The single inoculations were designed to determine if a particular fungal species was more virulent than the other, while co-inoculations, which only followed seedling treatment with Alamo, were used to evaluate fungicidal effects when fungi occurred in unison, as they would in the natural environment. Inoculations were carried out by first cleaning the outer bark with cotton rolls soaked in Clean-jet (25% isopropanol, 0.5% polyalkoxylated alcohol, Arborjet Inc., Woburn, Massachusetts, U.S.). A sterile razor blade was then used to make one slanting incision (~0.5 cm) into the stem. One full loop of inoculum was transferred from a one-week old fungal colony to each treated seedling using a flame-sterilized microbiological loop. After inoculation, the incision was closed and tightly sealed with Parafilm and aluminum foil to prevent contamination and desiccation and to immobilize the stem at the point of incision. For sterile agar and water controls, one loop full of sterile MEA or 100 μL of sterile water, respectively, were placed into the stem incision and similarly sealed. The seedlings were kept in a greenhouse under 400 watt metal halide lamps (10-hour light cycle) and watered weekly. Assignment of seedlings to treatments and location of seedlings in the greenhouse was according to a completely randomized design. Study seedlings were assessed weekly for foliar necrosis and destructively autopsied by dissection after 120 days to document lesion development and to re-isolate fungal species from cross sections of seedling stems.
Phytotoxicity of fungicides
Another 40 seedlings were randomly assigned to four treatments to test the phytotoxicity of injected TREE-äge, Alamo, and Arbotect 20-S, relative to an untreated control (UTC). A Micro I.V. designed for this experiment (Arborjet, Inc., Woburn, Massachusetts, U.S.), based on a 10 cc capacity syringe fitted with 18 ga × 2.5 cm syringe needle, administered TREE-äge, Alamo, or Arbotect by gravity feed into P. monticola seedlings. All treatments were infused by drip, where each seedling received 5 mL of a 10% solution. Injections were conducted by first cleaning the injection site using cotton rolls soaked in Clean-jet. A sterile razor blade was used to make a thin incision into the sapwood of the seedling, just large enough to allow a tight fit of the syringe tip. The syringe was then placed into the incision, while elastic horticultural tape was used to secure the site. The valve on the Micro I.V. was opened to allow the pesticide to be administered by slow drip. The Micro I.V. was not removed until all of the solution had drained. The seedlings were arranged randomly in a separate section of the greenhouse from that which was occupied by the pathogenicity study, and so were held under the same environmental conditions previously described. Seedlings were visually inspected weekly for foliar phytotoxicity (e.g., foliar necrosis) and destructively autopsied after 120 days to document vascular health.
Autopsies and statistics
Seedlings were observed weekly to note symptoms of foliar necrosis and autopsies were conducted at 120 days post-treatment, earlier if all needles were dead, to assess cause of death (Table 1). Where oxidized or necrotic phloem completely encircled the stem, the seedling was categorized as dead. The earliest mortality was observed at 40 days in seedlings inoculated with G. clavigera. After 120 days, the percent circumferential lesion was measured in all remaining treatments. Circumferential lesion formation reflects the extent of stem girdling, a critical metric to tree survival. Autopsies were conducted using the following steps: 1) the bark was removed at the base of each seedling, up to the first node (first year stem growth); lesion circumference and height were measured; and 2) thin transverse sections of treatments were taken for microscopic examination of discoloration (oxidation) and occlusions of the vascular tissues.
Evaluation of mortality 120 days after inoculations and injection of Pinus monticola seedlings with respective treatments in Experiment 1. Mortality rates for each treatment are based on 10 seedlings.
Statistical analyses were carried out separately for each experiment. For each experiment, the null hypothesis that mortality was the same for all treatments was tested using Fisher’s exact test (FET) on an Rx2 frequency table with seedlings classified as living or dead for each of R treatments (R = 5 or 4 for the pathogenicity and phytotoxicity studies, respectively). Rejection of the null hypothesis at significance level 0.05 was followed by pairwise comparisons of treatments using FET with Bonferroni-adjusted P-values on the corresponding 2x2 tables. For the pathogenicity study, the two control treatments were compared first and, when a non-significant outcome (P > 0.10) was observed; they were pooled for comparisons with other treatments (Table 1). One-way analysis of variance (ANOVA) was used to compare the differences in foliar browning, circumferential girdling, and axial lesion length. Lesion length was log-transformed prior to analysis to decrease variance heterogeneity. Pairwise comparisons among treatments were carried out with Tukey’s HSD procedure at the 0.05 level of significance using the MIXED procedure in SAS.
Experiment #2
Dual inoculation of G. clavigera
As before, P. monticola seedlings (mean caliper and height of 0.5 ± 0.02 and 15.0 cm ± 0.4 SE respectively), were purchased from the University of Idaho, Franklin H. Pitkin Forest Nursery (Moscow, Idaho, U.S.). In this experiment, G. clavigera and sterile MEA agar, with 10 replicates per treatment were employed. Leptographium longiclavatum was not used in this experiment because it failed to survive cold storage between experiments. Inoculations were made as in Experiment 1, but with a second inoculation being made on the opposite side of the stem (180 degrees to the first) and offset by 1 cm in height. The lesion data from Experiment 1 were used to determine the offset distance between incisions in this experiment. Seedlings were kept in a greenhouse under the same environmental conditions as previously stated and destructively autopsied after 40 days. This was sufficient time to observe needle browning. Lesion development was not documented, as researchers were only concerned with tree mortality. Cross sections of stems with fungal inoculations were plated on MEA to recover fungal species.
RESULTS
Fungicide Screening
The effect of concentration on repression of G. clavigera growth in vitro (Table 2) was not the same for all fungicides (F8,29 = 17.76, P < 0.001). There was no evidence of repression with increasing concentrations of TREE-äge (from 1 to 10,000 ppm, t31 = 0.05, P = 0.962). In contrast, there was significant repression observed with increasing concentrations of the fungicide treatments Alamo and Arbotect (t31 = 15.46, P = < 0.001 and t31 = 10.28, P = < 0.001, respectively). Treatments of Alamo at 1,000 and 100 ppm, showed significantly greater repression than respective concentrations of Arbotect (P < 0.05) (Table 2). However, Alamo at 10 and 1 ppm, and Arbotect at 100, 10, and 1 ppm, were not significantly different in repression than the control (P > 0.05) (Table 2).
In vitro screening of pesticides TREE-äge, Alamo, and Arbotect, measured as repression in mm distance from G. clavigera colony. Means and standard errors (SEs) are based on two treated discs in each of three dishes and result from an ANOVA with nested errors.
Experiment #1
Pathogenicity of fungal isolates
After inoculations, L. longiclavatum and G. clavigera successfully colonized the sapwood and resulted in mortality of 50 and 90% of seedlings, respectively (Table 1). One seedling in the DI water control group (10% mortality) and two in the sterile agar treatment group (20% mortality) died. As mentioned, these two control groups were pooled (15% mortality) for statistical comparisons, resulting in a significantly lower mortality than that observed with G. clavigera (FET, P < 0.001) but no difference in that observed with L. longiclavatum (P = 0.467) (Table 1). Ten percent mortality was observed in seedlings injected with Alamo and co-inoculated with L. longiclavatum and G. clavigera. This was compared to 50% and 90% in L. longiclavatum and G. clavigera, respectively, the reduction in mortality being significant when compared to that for seedlings inoculated with G. clavigera (P = 0.007) but not L. longiclavatum (P > 0.05). Observed percent foliage necrosis was 56.7 ± 14.5 and 91.0% ± 9.0 SE in the L. longiclavatum and G. clavigera-inoculated seedlings compared to 20.8% ± 9.2 SE in the Alamo/blue-stain inoculation (Table 3) and 20.5% ± 7.9 SE in the pooled controls (DI water), respectively. Percent foliar necrosis was significantly higher for seedlings inoculated with G. clavigera than for Alamo/blue-stain-inoculated seedlings and the pooled controls (Table 3). Circumferential girdling was calculated by measuring lesion width in centimeters and dividing it by stem circumference. Lesions averaged 92.0 ± 5.7, 51.8 ± 7.4, 38.7 ± 5.7, and 28.9% ± 6.4 SE of stem circumference for G. clavigera, L. longiclavatum, Alamo/blue-stain inoculation, and inoculated checks combined, respectively (Table 4). Girdling was significantly greater for seedlings inoculated with G. clavigera than for the other inoculation treatments and the pooled controls. Axial lesion lengths were greatest in G. clavigera–inoculated seedlings, followed by L. longiclavatum, Alamo/blue-stain, and controls (4.8 ± 0.5, 2.3 ± 0.4, 0.7 ± 0.1, and 0.6 cm ± 0.1 SE, respectively) (Table 4). Dissections revealed that G. clavigera and L. longiclavatum inoculations had oxidized and brownish-blue discolored resin ducts, parenchyma ray cells, and phloem, while the sterile agar and wa ter controls exhibited clear, light-colored sapwood and healthy phloem (Figure 1). The Alamo/blue-stain-inoculated seedlings also showed clear-light sapwood and healthy phloem, except for the dead seedlings, which exhibited dark vascular lesions matching those observed from G. clavigera and L. longiclavatum. Cross sections above and within the inoculation site of the controls indicated successful compartmentalization of incision wounds. Grossmannia clavigera and L. longiclavatum, identified by morphological analysis, were recovered from the excised cross sections.
Percent foliar dieback in pathogenicity and phytotoxicity studies; means and standard errors (SEs) for each treatment are based on 10 seedlings, except for the inoculation controls (where the average of water and sterile agar treatment means and appropriate SE are reported).
Pathogenicity of inoculation treatments as measured by percent stem girdling and lesion length (cm).
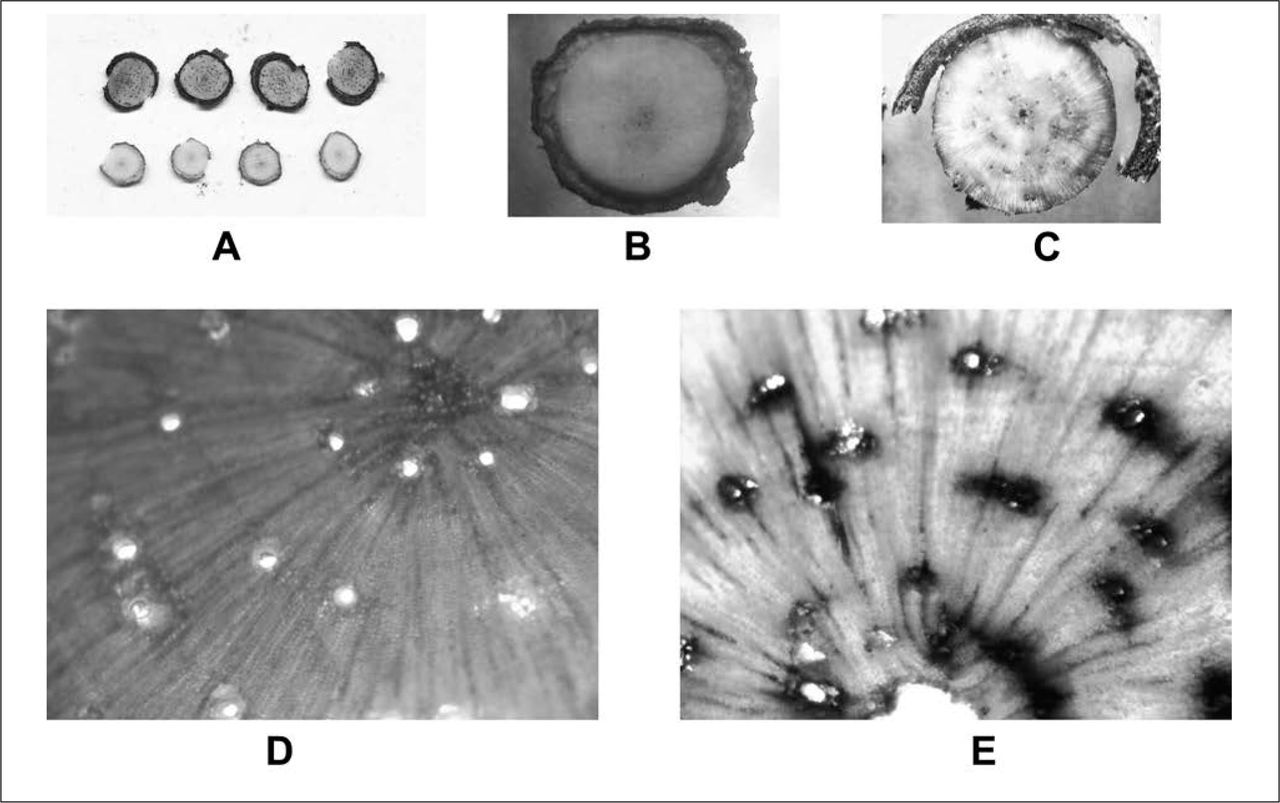
Cross-sectional destructive autopsies of P. monticola seedlings conducted at 120 days: A) Comparison of a subset of blue-stain-fungi-inoculated seedlings (top, discolored) versus untreated controls (bottom, healthy). B) Water-inoculated seedling with healthy and intact vascular (xylem and phloem) tissues (40x). C) Grosmannia clavigera-inoculated seedling with discolored vascular tissues and necrotic phloem (40x). D) Clear, healthy sapwood from sterile agar-inoculated seedling (100x). E) Leptographium longiclavatum-inoculated seedlings with oxidized and discolored resin ducts and radial parenchyma (100x).
Phytotoxicity of fungicides
After 120 days, mortality of seedlings treated with TREE-äge, Alamo, Arbotect, or untreated checks was 0%, 0%, 0%, and 10% mortality, respectively (Table 1), giving no indication of differences among pesticide treatments and control (FET, P = 1.0). Foliage necrosis did not differ significantly (F3,36 = 1.98, P = 0.135) among the untreated controls, TREE-äge, Alamo, and Arbotect treatments, with 30.2 ± 8.5, 13.5 ± 1.8, 19.6 ± 3.2, and 25.0% ± 4.2 SE, respectively (Table 3). No foliar phytotoxicity was therefore observed due to application of any systemic chemistry at the rates used. Microscopic examination confirmed that discolored sapwood was present and limited in the TREE-äge, Alamo, and Arbotect treatments; resin ducts and phloem were neither oxidized nor necrotic. Cross sections of a single untreated control seedling that died had oxidized xylem and an unidentified infection, while all other untreated controls had clear sapwood and healthy phloem.
Experiment #2
Dual inoculation of G. clavigera
Inoculations of G. clavigera on both sides of seedling stems resulted in 91.0% ± 9.0 SE seedling mortality, compared to 0.0% of the sterile agar controls. G. clavigera was identified by morphological characteristics and was re-isolated from cross sections of inoculated seedlings.
DISCUSSION & CONCLUSIONS
The two Ophiostomatoid fungi isolated from western white pine, Grosmannia clavigera and Leptographium longiclavatum, caused mortality of P. monticola seedlings following inoculation. Seedlings faded as early as 40 days following inoculation with G. clavigera, whereas seedlings generally faded more slowly following inoculation with L. longiclavatum. These results may differ in mature trees, as trials were only performed on seedlings. However, Krokene and Solheim (1998) demonstrated that the results of inoculations of two- and four-year-old Norway spruce seedlings, with four bark beetle-associated blue-stain fungi, largely agreed with previous results of inoculated 40-year-old Norway spruce trees with the same fungal strains. These, as well as the current results, suggest that inoculation of seedlings can be a reliable bioassay to predict the pathogenicity of blue-stain fungi associated with bark beetles. As L. longiclavatum is a recently described species, information on its virulence is not extensive. Lee et al. (2006b) proved its pathogenicity on 98- to 130-year-old lodgepole pine (P. contorta), suggesting that it may contribute to the mortality of MPB-infested pines. Rice et al. (2007b) compared the virulence of the three main associated blue-stain fungi—Ophiostoma montium, G. clavigera, and L. longiclavatum—in lodgepole pine, jack pine, and lodgepole × jack hybrids, concluding that these species are about equally competitive and virulent, finding no significant differences in lesion lengths between G. clavigera and L. longiclavatum. The results of the current study differ from this, as a significant difference was observed in mortality and extent of circumferential lesions over time between the two species, with G. clavigera being more virulent. This difference could be due to the variability of virulence in geographically distant strains of G. clavigera and L. longiclavatum, attributed to host-specific effects (western white pine versus jack and lodgepole pines and their hybrids), or could also be attributed to the amount of inoculum used for infection. Further research including the addition of a second inoculation point on the same seedling with L. longiclavatum, may be helpful in elucidating this question.
This injection experiment used a newly developed technique, Micro I.V.’s, to inject the pesticide treatments, and from researchers’ observations it performed effectively. Of the 5 ml of TREE-äge, Alamo, and Arbotect placed into the Micro I.V.’s precise uptake into each seedling was not calculated. However, results indicate that the volume of solution administered and infused into the xylem tissues was sufficient to show effects in the xylem (e.g., lesion length). Seedling autopsies also revealed discoloration from dye, a component in the TREE-äge and Alamo formulations, visually confirming that the vascular tissues were exposed to the injected product. Seedling mortality was not observed following injection with TREE-äge, therefore, the method of injection and application of TREE-äge per se is not likely to cause injury to western white pines.
Since the isolates were obtained from TREE-äge-injected trees attacked by MPB, researchers strongly suspect the involvement of G. clavigera and L. longiclavatum and other MPB associates in tree mortality. Furthermore, it was demonstrated that G. clavigera and L. longiclavatum are pathogenic with differences in virulence on P. monticola seedlings. Field studies to evaluate tree protection using TREE-äge and Alamo in P. monticola trees attacked by MPB are currently underway, and will help determine the generality of these findings, and, more specifically, provide an indication of their relative importance in determining mortality of larger trees in situ.
In vitro studies provide useful information in the development of effective injection treatments for tree protection. These results demonstrate that TREE-äge alone did not provide any repression of G. clavigera, and are consistent with the recovery of isolates of G. clavigera and L. longiclavatum from treated trees. Repression of the fungi was evident in both Alamo and Arbotect injections, particularly at the higher concentrations (Table 2), suggesting that similar chemistries could be used for systemic treatment against blue-stain fungi. Research utilizing injection of both insecticide and fungicide chemistries is currently being conducted to evaluate protection against MPB/blue-stain associates in situ with P. monticola, the results of which will help to determine efficacy in tree protection. In the interim, it is advised that tree stewards and arborists consider both vector and microbial associates when treating trees with systemic pesticides.
Acknowledgments
This study was made possible through the support of the USDA FS, Southern Research Station, RWU-4552, Pineville, Louisiana, U.S.; Region 5 Forest Health Protection, Susanville, California, U.S.; and the Modoc National Forest, Alturas, California, U.S. The authors thank Cavell Brownie, Professor Emeritus, North Carolina State University, Raleigh North Carolina, U.S. for conducting the statistical analyses in this study. The authors also thank David Cox, Syngenta Crop Protection, LLC. for his review and comments. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the United States government.
- Copyright © 2016, International Society of Arboriculture. All rights reserved.









{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.