
Abstract
One codominant stem on each of 48 similar Quercus virginiana Highrise® trees was pruned to evaluate impact of pruning severity on growth suppression and partitioning. Targeted pruning severity (0, 25, 50 or 75% foliage and subtending branches removed) based on visual estimates of two people correlated well (r2 = 0.87) with the ratio stem cross-sectional area removed: cross-sectional area at base of the pruned codominant stem. Pruning reduced cross-sectional area growth on codominant stems compared to the leader stem that was not pruned, especially during the first 12 months following pruning. Increased pruning severity reduced cross-sectional area growth on the pruned stem in proportion to amount of foliage removed. In each of three years following pruning, cross-sectional area of the unpruned leader stem increased more on trees receiving targeted pruning severities of 25% or 50% than trees pruned with the 75% severity or trees not pruned. Shift in growth from the pruned to unpruned portion of the tree reduced diameter ratio between the two stems, which should make the union stronger. Diameter ratio changed most for the 75% pruning severity.
Branches are well-attached to the tree when they are small in comparison to the trunk (Gilman 2003; Kane et al. 2008). This results from shade suppressing branch growth in the forest encouraging growth in a dominant trunk. Codominant stems on trees result in a weak union (Kane et al. 2008), especially when accompanied with bark inclusions (Smiley 2003). However, there is little research on pruning strategies for urban trees such as growth suppression on codominant stems. Most pruning studies were conducted in forested stands, not on open grown landscape trees.
Rom and Ferree (1985) reported leaf, shoot, root, and total dry weight in the year after pruning peach in an orchard decreased as pruning severity (dose) increased. Stem diameter growth generally slows with increased crown raising severity (crown raising pruning type as described in American National Standards Institute 2008), and the effect in the forest can last two or more years (Langstrom and Hellqvist 1991; O’Hara 1991). In contrast, Pinkard and Beadle (1998) reported increased cross-sectional area growth in tops of forest trees with increased crown raising severity. There is little research on growth impacts of raising or any other pruning type on open-grown landscape trees. It is not clear whether raising conducted in the forest relates well to open grown trees common in arboriculture, but it is one of the only guidelines available in the literature.
Pruning did not alter trunk diameter or tree height of rose gum (Eucalytpus grandis W. Hill ex Maiden) (Bredenkamp et al. 1980), nor crown volume of black walnut (Juglans nigra L.) (Funk 1979). Neilsen and Pinkard (2003) showed that light crown raising (removing all branches in the lower 45% of tree height) had no effect on growth of Monterey pine (Pinus radiata) D. Don, but heavier pruning (60% or 75% removal) decreased trunk diameter growth, stem volume, and tree height in the forest.
Clark (1955) and Funk (1979) found that increasing amount of lower branches removed corresponded to reduced trunk taper of black walnut and more sprouting along the trunk. In contrast, some researchers found that neither stem taper (Pinkard and Beadle 1998) nor total timber yield (Bredenkamp et al. 1980) was affected by crown raising.
Pruning one side of the crown to encourage the other side to grow faster has been referred to as structural pruning (Gilman and Lilly 2008), but there is little research supporting this contention. One study on seedlings grown in an open landscape on California coastal live oak (Quercus agrifolia N´ee) and valley oak (Q. lobata N´ee) found that headed or thinned codominant stems grew slower than stems that were not pruned (Downer et al. 1994). Structural pruning seeks to enhance growth on the unpruned leader by reducing growth rate on pruned low branches and on pruned codominant stems higher in the crown. The pruning severity required to cause this growth reduction has not been thoroughly studied.
The objective of the current study was to determine impact of pruning severity on tree growth partitioning over time.
MATERIALS AND METHODS
Quercus virginiana Mill. is a common tree in USDA hardiness zones 7–10, and readily develops codominant stems, making the tree a good subject for study. In May 2005, forty-eight 11 cm (4.4 in) trunk caliper (SD 1 cm), 7 m (23 ft) tall (SD 48 cm), seven-year-old Quercus virginiana Mill. Highrise® live oaks planted three years earlier and 4.9 m (16 ft) apart were pruned to remove branch biomass of one codominant stem. All trees were surface-fertilized in a 3.7 m (12 ft) x 4.9 m (16 ft) area centered on the trunk with 1 kg (2.4 lbs) of 16:4:8 (N:P:K) one or two times a year between 2003 and 2007. On each tree, the two largest and similarly sized codominant stems growing from the same union were located, and a diameter tape was used to measure diameter at the base of each stem, 5 cm (2 in) beyond any swelling associated with the union. The smaller of the codominant stems (“the codominant”) averaged 47 mm (1.9 in) in diameter (SD = 13 mm), and was pruned according to the targeted severity. The larger of the codominant stems (“the leader”) averaged 62 mm (2.4 in) in diameter (SD = 16 mm), and was not pruned. The smaller stem was chosen for pruning to eliminate any possible effect of diameter ratio on growth response. Both stems were marked close to the union so they could be measured in subsequent years at the same position.
Pruning was applied by removing branches on the pruned codominant stem as one of four visual targeted pruning severities (TPS): 0% (control), 25%, 50%, or 75% of total foliage removed. Severities were visual estimates of the percentage of foliage removed from the pruned codominant stem. Visual estimates were made by two assessors standing next to the tree as stems were removed. Both assessors had to agree on the severity before pruning was considered complete. To quantify amount of biomass removed, a cross-sectional area (CSA) of each pruning cut (outside bark) was calculated using a diameter tape and summed for each pruned tree. Ratio of CSA removed to CSA at base of the codominant was termed CSAR. Pruning severities were arranged in a randomized complete block design with 12 blocks and four trees per block. One tree in each block randomly received one of four severities.
One to five pruning cuts (mean 2.9) were made on each pruned stem to attain the TPS; most cuts were reduction cuts and some were removal cuts (Gilman and Lilly 2008). The largest diameter, most upright portion of the codominant stem closest to the unpruned leader was removed first; others were removed, typically toward the end of the codominant stem, as needed to attain targeted severity. This structural pruning type is described in detail in Gilman and Lilly (2008). In September 2006, June 2007, and May 2008, diameters of pruned codominant stem and unpruned leader on each tree were measured in the same position as described above.
The relationship between branch or stem diameter and fresh foliage mass was calculated by choosing three trees at random from the source nursery field. Six branch and 6 stem segments were paired (for a total of 12) by equal diameter for the following diameters: 0.8 cm (0.3 in), 1.5 cm (0.6 in), 2.3 cm (0.9 in), 3.0 cm (1.2 in), 3.8 cm (1.5 in), and 4.6 cm (1.8 in); these represented orthogonal values within the range of pruning cut diameters. Diameter was measured with a diameter tape at the base of each branch or stem 5 cm beyond any swelling associated with the union. Branches were less than one-third diameter of trunk measured just above the union, and stems were at least three times branch diameter at the union. All leaves were carefully pulled from the segment distal to the diameter measurement and weighed fresh. Fresh weight was used because arborists work with live trees. A least squares regression line was fitted to stem or branch diameter and foliar fresh weight separately, then data combined for stems and branches because slopes of both relationships were statistically similar (t-test, P > 0.05; slope and intercept P > 0.05).
The percent increase in CSA growth of the codominant stem was calculated as [(CSA end of project – CSA at initial pruning) ÷ CSA at initial pruning] x 100. Each year, CSA growth of the pruned codominant stem and unpruned leader stem was calculated as CSA current – CSA previous measurement. Stem diameter was measured with a diameter tape at the base of each pruned and unpruned stem 5 cm beyond any swelling associated with the union. Trunk diameters were measured at the beginning of project and annually with a diameter tape at 30 cm (12 in) from ground.
CSAR was regressed onto TPS. Fresh leaf mass was regressed onto CSA of branches from which leaves were removed. The percent increase in CSA growth of the codominant stem was regressed onto CSAR. Linear relationships were analyzed with Minitab version 14 (Minitab Inc., State College, PA) and considered significant at P < 0.05. CSA on the codominant and leader, and ratio of diameter of the codominant to the diameter of the leader stem were analyzed by repeated measures two-way analysis of variance within a randomized complete block design using GLM in SAS 9.1.3 (SAS Institute, Cary, NC) with P < 0.05. Main effects were pruning severity and time. Trunk diameter was analyzed by repeated measures oneway analysis of variance with pruning severity as the main effect. Means were compared using Tukey’s multiple range test.
RESULTS
The number of pruning cuts required to achieve TPS increased with pruning severity from 2.7 for 25% severity to 2.9 for 50% severity, and 3.6 for 75% severity. Pruning cut diameter ranged from 8 mm (0.3 in) to 48 mm (1.9 in) across trees in all treatments.
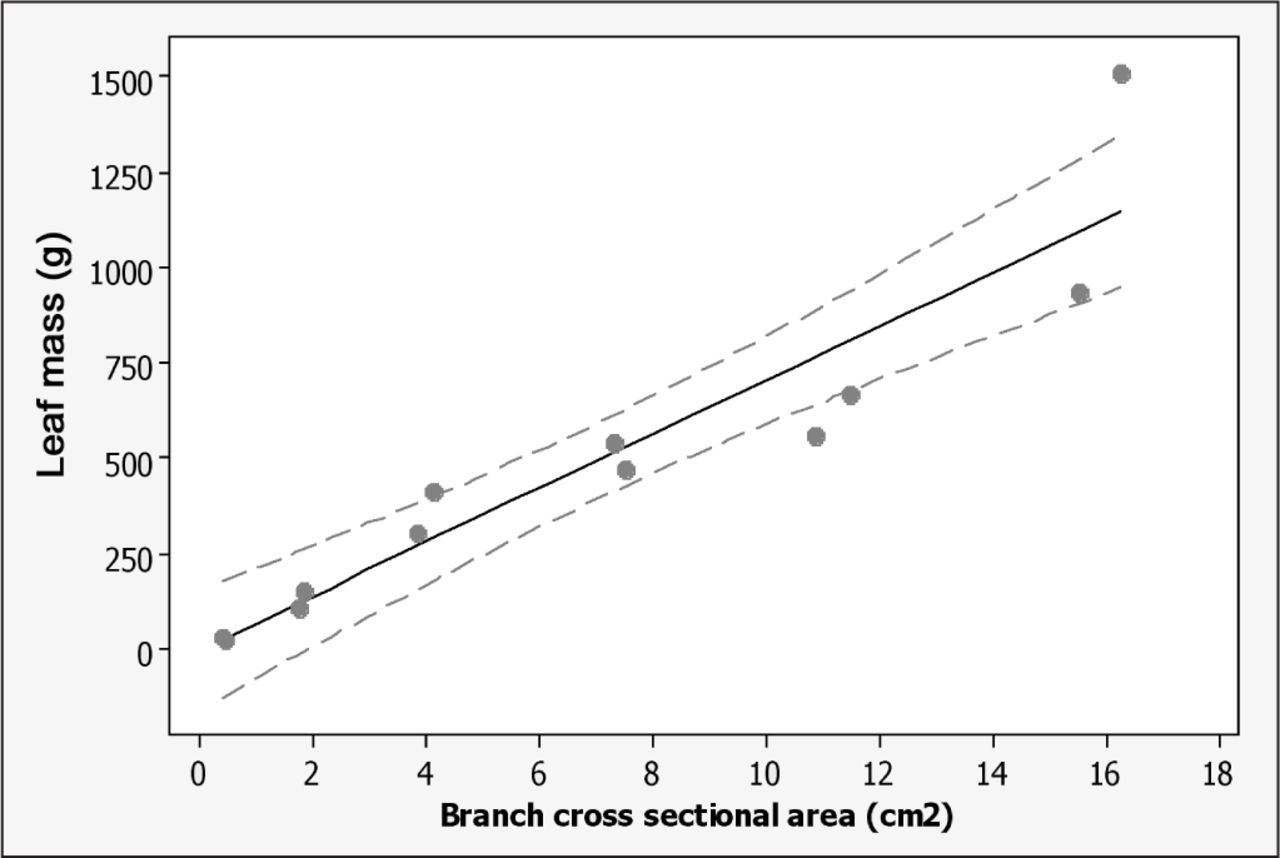
CSAR correlated well with TPS but varied widely within each TPS (Figure 1). This large range is attributed to variation in the visual estimate of foliage removed because there was an excellent relationship between CSA removed and actual foliage mass removed (Figure 2). Since slope in Figure 1 was close to one (0.99), mass of foliage removed from a live oak stem might be predictable visually by a trained assessor, although Smiley and Kane (2006) also found considerable variability in visual estimates. The CSAR of one pruned stem exceeded the CSA of the stem base and on two other pruned stems was close to 100% CSAR (Figure 3). Since these were all 75% pruning severity treatments, there was a visual 25% of foliage remaining on these three stems.

Relationship between percentage of cross-sectional area removed on the codominant (CSAR) and targeted pruning severity (TPS). CSAR = 0.99 (TPS) + 8.18. r2 = 0.69, slope P < 0.001, intercept P = 0.19. The dashed lines represent a 95% confidence interval for the regression equation.

Relationship between fresh leaf mass and branch cross-sectional area (CSA). Leaf mass = 70.81 (CSA) – 5.9, r2 = 0.88, slope P < 0.001, intercept P = 0.93. The dotted lines represent a 95% confidence interval for the regression equation.
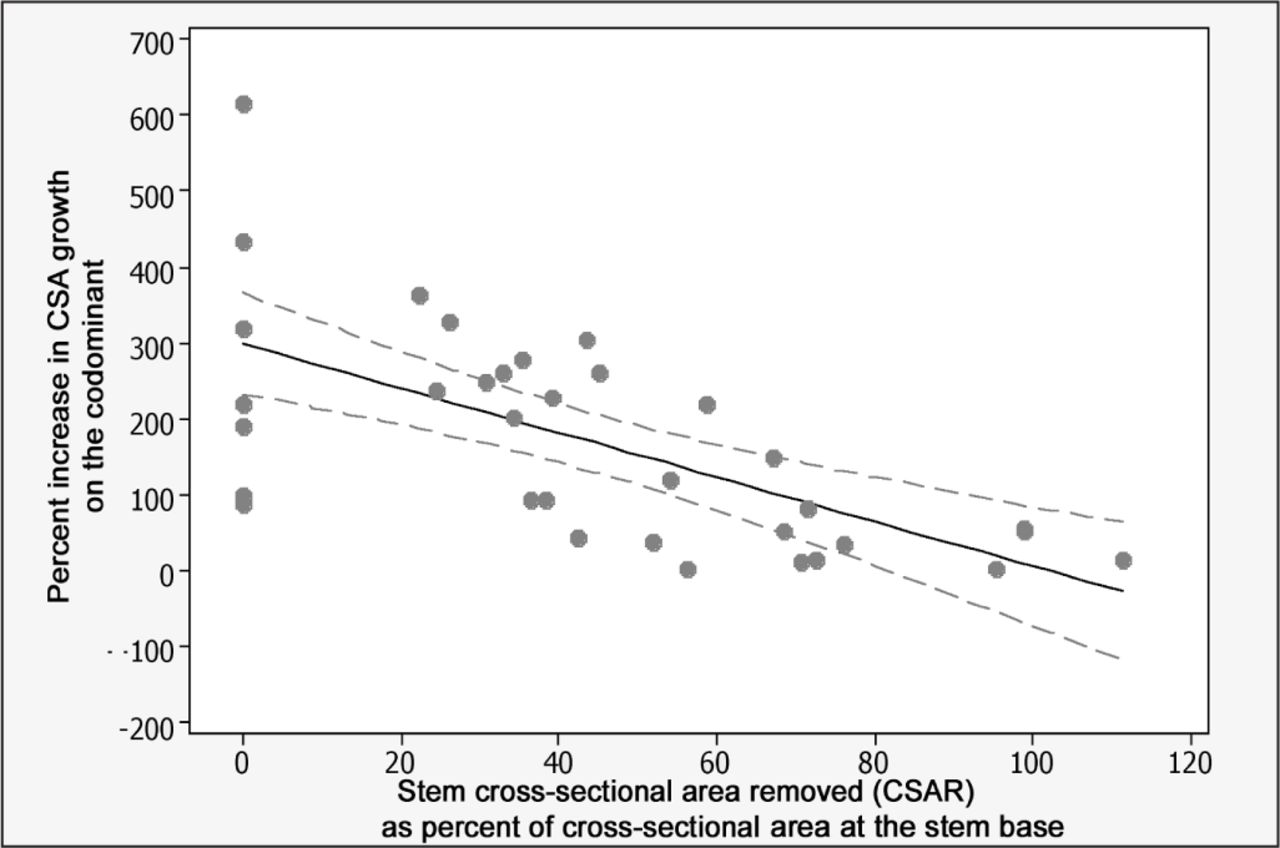
The percent increase in CSA of the codominant between May 2005 and May 2008 following removal of increasing amounts of stem cross-sectional area (CSAR). Percent increase = 298.7 − 2.93 (CSAR), r2 = 0.43, slope and intercept P < 0.001. The dotted lines represent a 95% confidence interval for the regression equation.
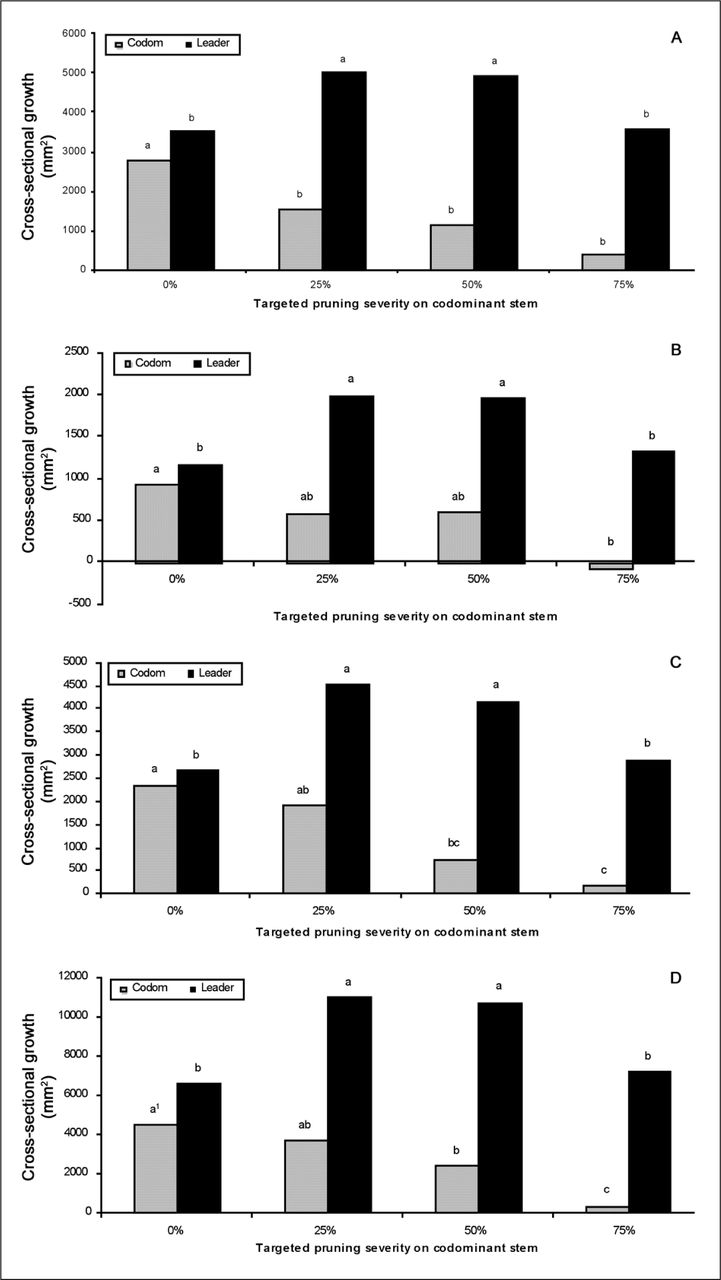
Increasing pruning severity slowed percent increase in CSA growth in proportion to amount of foliage removed on the pruned codominant stem (Figure 3). CSA growth on pruned codominant stems at 25, 50, and 75% TPS was less than on codominant stems on unpruned control trees in the first growing season after pruning (Figure 4A). This indicated that pruned codominant stems grew more slowly than codominant stems that were not pruned. CSA growth on pruned codominant stems was not affected at 25% or 50% pruning severities in the second year following pruning compared to unpruned trees, but stems receiving 75% severity grew slower than unpruned trees (Figure 4B). The 50% and 75% severities both grew more slowly than unpruned controls in the third year after pruning (Figure 4C).

Cross-sectional area growth of codominant and leader stem following removal of targeted pruning severity (0, 25, 50, and 75%): A) one, B) two, and C) three years after pruning, and D) combined data for all three years. Note: Within a stem type (codominant or leader), bars with the same letter are not statistically different at P < 0.05. Codominant stems are not compared to leader stems.
Pruning the codominant enhanced CSA growth in the unpruned leader (compared to the leader stem in unpruned trees) on trees receiving 25% and 50% pruning severities in all three years (Figure 4A-D). Resources appeared to be preferentially allocated to the unpruned leader at these lower pruning severities. This shift in growth from the pruned to unpruned portion of the tree reduced the diameter ratio of codominant to the leader for all TPS, except unpruned trees (Figure 5). Most of the reduction in diameter ratio occurred in the first year after pruning, and diameter ratio for 25% and 50% TPS remained unchanged in years two and three following pruning, which is reflected in the interaction between TPS and year. Trees that received 25% TPS on the codominant grew more in trunk diameter (caliper) beginning in the first year following pruning than trees that were not pruned and trees that were pruned more severely (Figure 6).
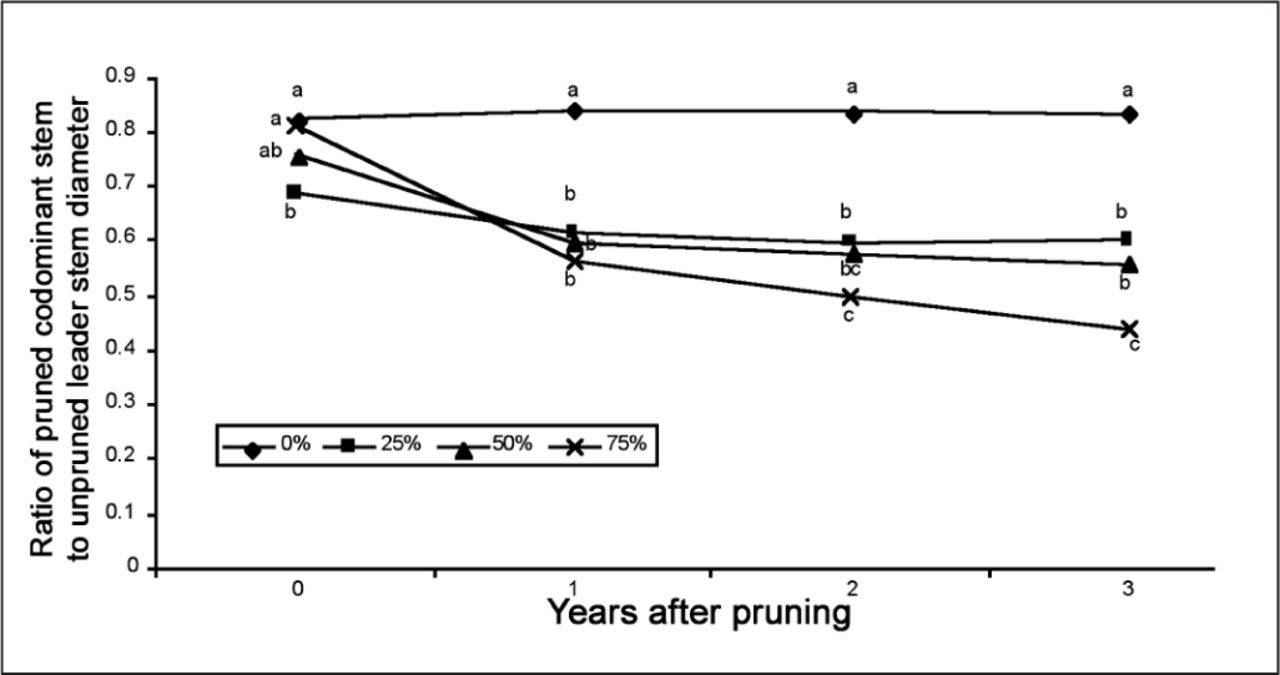
Diameter ratio of codominant to leader measured at the base of each stem at pruning (year 0) and in three subsequent years for four pruning severities: 0, 25, 50 and 75% TPS. Within each year, numbers followed by the same letter are not statistically different at P < 0.05.
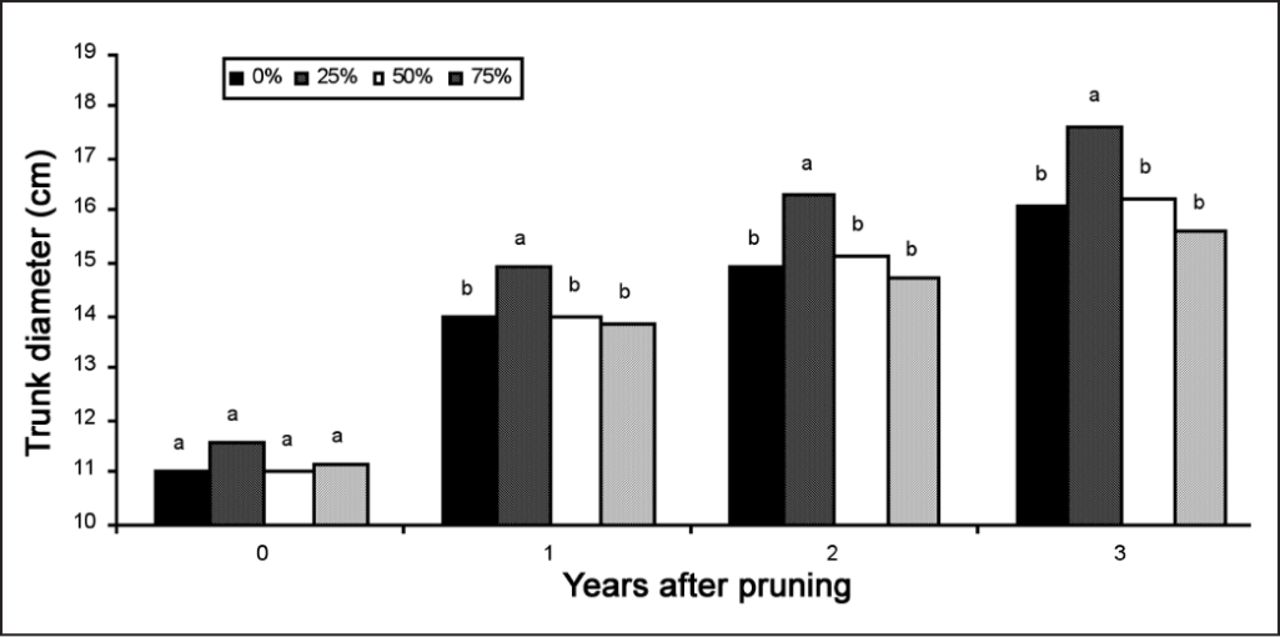
Trunk diameter 30 cm (12 in) from ground at pruning and in three subsequent years for four pruning severities (0, 25, 50 and 75%) from one codominant stem. Note: Within a year, bars with the same letter are not statistically different at P < 0.05.
Pruning the codominant with 75% severity did not affect CSA growth in the unpruned leader (compared to the leader stem in unpruned trees) (Figure 4). However, diameter stem ratio in trees that received 75% TPS continued to decrease with time so that by the third year it was significantly lower than in any other treatment (Figure 5). Change in diameter ratio was larger for larger pruning severities.
DISCUSSION
Net dry mass assimilation rate was not impacted by light lateral branch shortening (28% foliage and subtending branches removed) on single leader three-year-old alder (Alnus glutinosa L. Gaerten) (Singh and Thompson 1995). In contrast, Rom and Ferree (1985) showed that photosynthesis in single leader one-year-old peach increased for more than four weeks following light pruning (main stem headed) which could have caused the slight but statistically significant trunk growth increase in the present study (Figure 6). Although Stein (1955) and Clark (1955) also found trunk diameter enhancement with light pruning, research on many forest-grown conifers shows either a small reduction or no impact on trunk diameter growth from light crown raising (Hanley et al. 1995). Rate of trunk diameter growth in the present study was not reduced despite removing up to 75% of the foliage from the codominant stem. This might be due to the less than 30% (estimated) of total foliage on the tree removed with 75% TPS because only one stem on the tree was pruned. These were also young trees; stems of this diameter on older trees may have responded differently.
Since light pruning (25% TPS) shifted growth from pruned codominant stem to unpruned leader (Figure 4) and increased trunk caliper, this might appear to be a good method of encouraging development of a dominant leader in live oak trees without sacrificing trunk growth. However, growth shifted for only one year suggesting this amount of pruning might be most appropriate in situations where regular pruning can be conducted. Trees with medium pruning severity (50% TPS) also shifted growth to the unpruned leader, but unlike light pruning growth shifted for years one and three. Apparently, pruning the codominant stem removed enough resources to reduce wood accumulation in the pruned side while increasing accumulation on the unpruned side of the tree. The unpruned leader became the sink for increased allocation perhaps from the root system because of reduced demand and storage capacity on the codominant stem. Stored carbohydrates can be solubilized and translocated from roots to shoots in response to pruning (Singh and Thompson 1995). Pruning at 75% TPS dramatically reduced growth of the codominant as well as the diameter ratio of the codominant to the leader without affecting trunk caliper growth. This amount of pruning might be best suited for situations with longer pruning cycles or where dramatic growth reduction is warranted on codominant stems. Pruning with higher severities results in a large canopy void which could be aesthetically objectionable in some situations.
Increased pruning severity reduced growth on the codominant in proportion to amount of foliage removed. Langstron and Hellqvist (1991) found that removing 48%–75% of lower crown slowed growth for four years on Scots pine (Pinus sylvestris). The data showed the growth slowing effect using reduction and removal cuts lasted three years with the highest pruning severity. Few other studies have examined tree response to reduction pruning only one side of a crown other than Downer et al. (1994). Clearly this needs to be studied for a variety of tree types of different ages in different regions. The type of pruning cut (e.g., reduction versus removal), also needs to be evaluated for its impact on reduction in growth rate on pruned stems. Only Downer et al. (1994) compared pruning cut types, and showed that heading and removal cuts retarded growth similarly. Most studies done on forest trees have evaluated removal cuts from the lower and middle trunk, but not reduction cuts.
Gilman (2003), Kane et al. (2008), and others showed that smaller branch:trunk diameter ratios are associated with greater branch union strength. Eisner et al. (2002) also showed that branches with smaller ratios conduct less water per unit of a cross-sectional area. This might help partially explain how the branch protection zone functions by retarding movement of xylem inhabiting organisms such as fungi and bacteria. Structural pruning to induce a smaller stem diameter ratio as accomplished in this study at all pruning severities should make a tree structurally stronger, and perhaps more resistant to movement of decay organisms from branches to trunk.
Although there is inherent variability with using fresh leaf mass due to soil, climatic, weather, and species variation, regression coefficient (70.8, Figure 2) in the current study on Q. virginiana Highrise for branch CSA versus fresh leaf mass was similar to the 86.7 coefficient in Q. virginiana Cathedral Oak® (Grabosky et al. 2007). They showed that CSA on primary branches (those attached to the trunk) correlated well (r2 = 0.95) with actual fresh foliage mass on that branch, but the sum of CSA on the trunk above the union and adjacent primary branch was greater than CSA of trunk immediately below the union. This can be at least partially accounted for by hydraulic constriction represented by the branch protection zone found at the base of branches that are smaller than the parent branch (Eisner et al. 2002).
Pruning as performed in the current study created openings in the crown. Stem motion in windy weather was reduced when crown openings were large (Gilman et al. 2008), but openings created by pruning can close as branches change shape and move closer together in a windstorm if pruning severity is small (Vollsinger el at. 2005). Presumably (pers. obs.), openings close with time from a combination of growth from below on the subordinated stem and growth from above on the unpruned leader stem but how this happens has not been described. The current data provide evidence that growth-slowing effects on pruned codominant stems of live oak lasted three years at medium (50%) and high (75%) pruning severities in USDA hardiness zone 8b. Pruning with the lowest severity (25%) was not as effective in slowing growth on codominant stems. Pruning with a moderate to high severity appears to be a good method of reducing diameter ratio between pruned codominant stems and unpruned leader and should result in a stronger structure. Results may be different for different species and climates, and for larger trees.
CONCLUSION
Pruning reduced growth in linear proportion to amount of foliage removed on pruned codominant stems. This resulted in a reduction in size of the pruned codominant stem relative to the unpruned leader stem. Growth slowing effects from higher pruning severity lasted longer than for lower pruning severities. As much as 75% or more of a codominant stem can be removed without killing the stem on young live oaks pruned in this study. This provides guidelines for growers producing leaders when structurally pruning shade trees in a nursery, and for arborists pruning young trees in landscapes. It could also apply to younger outer portions of the crown of older trees where most structural pruning is conducted to resist storm damage.
Acknowledgments
Thank you to the TREE Fund and GreatSouthernTreeConference.org for funding of the project.

- © 2009, International Society of Arboriculture. All rights reserved.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.