
Abstract
Carbohydrate translocation, which follows anatomical and developmental patterns, is ruled by source-sink relations where energy-containing compounds are moved from sources of production to sinks of utilization. Seasonal carbohydrate concentrations in various tree parts were measured and compared in 10 cm (4 in) trunk diameter live oaks (Quercus virginiana P. Miller). Tissue samples from roots, trunks, twigs and leaves were collected from three-year-old field-grown trees on four dates throughout the 2005–2006 seasons. Laboratory analyses of glucose and starch were performed, and values were compared and contrasted according to sample location and time of year. Glucose levels were significantly higher in leaves during the winter (P ≤ 0.001), while starch concentrations were significantly higher in root and trunk tissues during the spring and winter assessments (P ≤ 0.001). Carbohydrate concentrations varied among tissues sampled within the tree. This study provides valuable information on the spatial and temporal partitioning of energy reserves, glucose and starch, in live oak so that arborists will have a better understanding of tree vitality, and the effects and environmental impacts of arboricultural treatments.
Carbohydrates are the principal products of photosynthetic activity and serve as the main energy reserve of trees (Tromp 1983). These products are used by organs where they are produced (e.g., leaves) or are translocated to other organs, a phenomenon controlled by sink-source relations (Allen et al. 2005). Therefore, carbohydrates can be translocated from organs that produce photosynthates (sources) to organs that produce little or no carbohydrates (sinks) where they can be used or stored (Taiz and Zeiger 2006). These sink-source relationships are influenced by tree vitality, nutritional state, environmental conditions, and the developmental stage of plants or tissues (Tschaplinski and Blake 1994; Grulke et al. 2001; Retzlaff et al. 2001). Understanding carbohydrate activity is critical in stressful environments, such as urban forests, where tree health is negatively impacted and environmental stressors are frequent.
Research has highlighted the important role of carbohydrate reserves on a tree’s ability to tolerate stressful conditions (Barbaroux et al. 2003). Nonstructural carbohydrates (i.e., starch and soluble sugars) influence the capacity in trees for supporting growth, metabolism, and ultimately their survival (Kaelke and Dawson 2005). Most of the research information about reserve translocation in trees has been explained for young plants grown in greenhouses or natural environments rather than for older trees where carbohydrate production, translocation, utilization, and storage may differ greatly (Tschaplinski and Blake 1994; Gansert and Sprick 1998; Tognetti and Johnson 1999; Domisch et al. 2002). Additionaly, much of the recent research concerning carbohydrate partitioning has focused on a single season or short period of time (DeLucia et al. 1998; Retzlaff et al. 2001; Barbaroux et al. 2003).
Trees vary in the allocation or use of carbohydrates stored in tissues. In deciduous trees, roots and trunk serve as the main storage organs during the dormant season, and their reserves are typically depleted shortly before leaves begin to emerge; conversely, evergreen trees store considerable amounts of starch in leaves and branches (Larcher 1980; Grulke et al. 2001; Retzlaff et al. 2001; Newell et al. 2002). Deciduous trees require extensive carbohydrate storage to maintain the living biomass and cope with stress-inducing factors (Abod and Webster 1991; Gansert and Sprick 1998). For example, white oak (Quercus alba L.) rapidly mobilizes and replaces starch reserves during the critical period of canopy generation in the spring (McLaughlin et al. 1980). Conifers accumulate carbohydrates in needles and twigs prior to bud-break and translocate them during the beginning of shoot growth (Ludovici et al. 2002). Some conifers such as Scots pine (Pinus sylvestris L.) allocate high percentages of sugar in needles as a response to low soil temperatures (Domisch et al. 2002).
Trees in urban environments are subjected to numerous environmental stressors throughout the year that negatively impact carbohydrate production, utilization, and storage. Live oak (Quercus virginiana) is a common species found in urban environments in the southern United States due to the species’ elegant canopy shape, adaptability to poor sites, low maintenance requirements, disease resistance, and long life span, which make this species suitable as an ornamental for urban environments (Little 1979; Gilman and Watson 1994). Because a live oak’s leaves continue to function throughout the winter until the trees defoliate during budbreak, the species is considered as semi-evergreen. Therefore, research on carbohydrate allocation will help to understand whether the species follows a deciduous or evergreen pattern. Even when under urban conditions, carbohydrate allocation can also be affected when tree organs are modified by human activities such as root pruning, canopy pruning or trunk damage (Harris et al. 2004).
Carbohydrate reserves help to offset low carbohydrate production due to stressful conditions or high demand. Differences in the allocation of carbohydrates to storage tissues could arise from varied requirements of different organs, different needs during growth, or maintenance respiration required among different species (Dean 2001; Barbaroux et al. 2003). Variation in starch concentrations could indicate different rates of production, demand, or shifts in allocation (Ludovici et al. 2002). In trees, the continuous pathways of transport and storage of previous-year assimilates are essential for subsequent growth processes, so the coordination and interrelations of morphogenic and photosynthetic processes are very important (Kaipiainen and Sofronova 2003). Carbohydrates can also be used under stress conditions as a precursor for secondary compounds used to resist biotic or abiotic stress (Webb 1981; Renaud and Mauffette 1991). Understanding carbohydrate partitioning in live oaks can help in understanding the translocation of the main carbohydrate, glucose and starch in semi-evergreen species as well as the relationships between phenological stages and movement and utilization of available energy resources.
The objective of this research was to study the nonstructural carbohydrate content, glucose and starch, in tree roots, trunks, twigs, and leaves of large, field-grown live oaks as well as to determine the impacts of seasonal influences on carbohydrate concentrations and utilization.
MATERIAL AND METHODS
Plant Material
Five, field-grown live oaks, about 10 cm (4 in) trunk diameter measured 30 cm (12 in) above ground, were randomly selected from within a nursery at Monaville, TX (29°57′1.59″N, 96°3′28.73″W). Trees selected in the 2005-year were planted in 1999 at 5 m (16.4 ft) spacing, and grown under similar conditions. The soil was a deep, moderately well drained, slowly permeable Lake Charles clay, and trees were annually fertilized (20-20-20, water soluble tree fertilizer).
Tissue samples from roots, trunk, and canopy were collected from opposing sides of the tree (across and between rows of trees) corresponding to the four cardinal points. Root samples consisted of 4 mm (0.16 in) diameter increment cores from the buttress (woody) roots [2 cm (0.79 in) from the base of the trunk]. Tissues from the trunk, cores 4 mm diameter, were collected using an increment hammer (Haglof©; Langsele, Sweden) at 1.3 m (4.3 ft) height from the ground. Increment cores were approximately 100 mm (3.94 in) in length. Canopy samples consisted of five 5 cm (2 in) long twigs with leaves, which were randomly collected from the lower two-thirds of the canopy (Mclaughlin et al. 1980). Samples from the different parts of the trees were collected within two hours in July 2005, September 2005, January 2006, and March 2006.
Samples were stored on blue ice (Rubbermaid®, Fairlawn, OH) immediately after collection in the field, transported to the lab within two hours, and oven-dried at 80 °C (176 °F) until weights stabilized (approximately 48 hours). After drying, samples were ground and stored in plastic bottles at −20 °C (−4 °F) until the carbohydrate concentrations were analyzed (Kolb and McCormick 1991).
Carbohydrate Analysis
Glucose and starch concentrations were determined for each sample using Sigma® GAGO-20 reagents (Sigma, St. Louis, MO). Glucose was extracted from tissues using methanol:chloroform:water (MCW, 12:5:3, v/v/v) solution, and 0.5 mL (0.02 fl oz) of the supernatant from the extract or glucose standards was mixed with 5 ml (0.17 fl oz) of anthrone reagent (Jaenicke and Thiong’o 1999). Absorbance of samples and standards were read at 625 nm within 30 minutes using a spectrophotometer (Spectronic 20, Baush & Lomb, Rochester, NY). Glucose concentrations were calculated through standard linear regressions and expressed as mg per g of dry weight. Starch content was determined by enzymatic conversion of starch to glucose. In the remaining pellet after glucose extractions, amyloglucosidase, an enzyme responsible for the conversion of starch to glucose, was used. Standards were prepared using potato starch, and the samples were read at 540 nm within 30 minutes. Starch content was calculated through standard linear regressions and expressed as mg of glucose per g of dry weight. (Haissig and Dickson 1979; Renaud and Mauffette 1991).
Data Analysis
An analysis of variance was conducted on data including in the model cardinal orientation, type of tissue, and time of year. Results from samples collected from opposite side of the tree (North-South, and West-East) were pooled in order to compare differences among and between rows in the nursery. The data was analyzed with the procedure General Linear Model (GLM) using the Statistical Package for the Social Sciences (SPSS, v. 13) for Windows (SPSS, Chicago, IL). Comparisons among tissues and within orientations were performed using Fisher’s Least Significant Differences (LSD) to determine differences in carbohydrate concentration in trees.
RESULTS AND DISCUSSION
Statistical analysis revealed no main or interactive effect (P > 0.5) of cardinal orientation. When data from opposing orientations (N-S and E-W) were pooled, no differences were found (data not shown). These results indicate that future research involving tissue sampling for carbohydrates may not be impacted by cardinal direction when sampling similar tissues at similar heights. However, carbohydrate reserve concentrations varied from base to the top in the trunk of sessile oak (Quercus petrea L.) and beech (Fagus sylvatica L.) (Babaroux et al. 2003).
Differences in glucose concentrations among tissues varied (P < 0.001) across seasons (Figure 1). Glucose content was higher in leaves than in other tissues tested through the year, except in September when there were no significant differences in glucose levels in leaves and roots. Although leaf glucose levels decreased in September, they were still higher than glucose levels in trunk and twigs. To the contrary, species such as pecan (Carya illinoinensis (Wangenh) K. Koch) or ponderosa pine (Pinus ponderosa Laws.) showed constant concentration of glucose during the growing season in different tissues of the tree (Grulke et al. 2001; Kim and Wetzstein 2005). Foliar glucose levels decreased in July and September, possibly due to the developmental stage of leaves (Figure 1). The glucose fraction in roots reached the highest concentration in the fall which may likely indicate a high translocation of sugars to roots for storage purposes. Root glucose levels were lower in the March–July period, which corresponds to the time when the roots had the highest levels of starch (Figure 2). The decrease in glucose content in leaves during fall can be a result of the high translocation and replacement of carbohydrates from source to sinks or storing organs (McLaughlin et al. 1980).

Glucose concentrations (mg/g dry weight) measured in live oak samples collected at four different dates. Bars represent the SE of the mean.
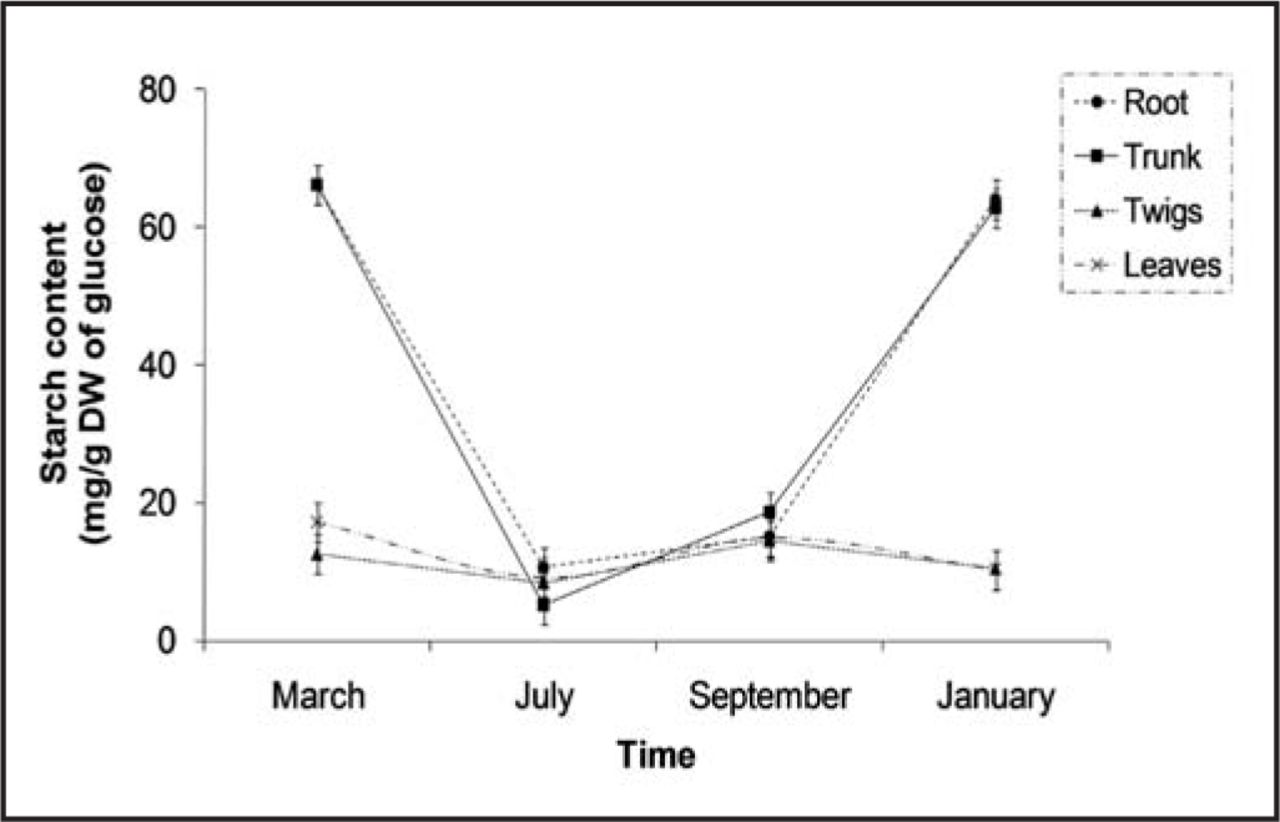
Starch concentrations (mg/g DW of glucose) measured in live oak samples collected at four different dates. Bars represent the SE of the mean.
Glucose concentrations in leaves might increase in winter because live oaks maintain live leaves throughout the winter as a response to environmental factors. Ludovici et al. (2002) also found a two-to-three-fold increase in winter glucose concentrations when compared to summer levels in needles of loblolly pine (Pinus taeda L). The high levels of glucose in leaves during the winter assessment can be caused as a response to the low temperatures [below 0 °C (32 °F)] that occurred before the sample collection. Conversion of starch to sugar is a physiological manifestation of cold hardiness in trees (Levitt 1980; Nguyen et al. 1990).
A significant interaction (P < 0.001) was also found for starch among time of year and tissues sampled. The highest starch concentrations were measured in roots and trunks during spring and winter (Figure 2), thus confirming their role in facilitating carbohydrate storage (Allen et al. 2005). Similar results were found for trunks and roots of sessile oak which had higher quantities of carbohydrate reserves in autumn than in late summer (Barbaroux et al. 2003). Trunk tissues showed a depletion of starch from spring to the summer (Figure 2). Gansert and Sprick (1998) identified starch disappearance during the summer in beech. Low concentrations of starch were also found during the spring in Scots pine (Domisch et al. 2002).
Starch concentrations found in roots and trunk (Figure 2) emphasize the importance of these two organs as carbohydrate reservoirs during the dormant season. These storage tissues have an indispensable role during the initiation of vegetative growth in the spring. The reduction of starch levels observed in the summer is likely a consequence of growth and high night temperatures and is associated with the hydrolyzation of stored carbohydrates for growth and maintenance (Kaipiainen and Sofronova 2003; Pallardy 2008).
Even when the interaction of type of tissue and time of year was significant, the annual mean carbohydrate concentrations varied among different tissues in trees (data not shown). Glucose levels in leaves were about double than those in twig, root, and trunk tissues. For starch, an inverse pattern was found where root and trunk tissues had more than three times the starch content than foliar tissues. This overall concentration pattern corroborates source-sink mechanisms for carbohydrate partitioning described in other studies (Kaipaiaien and Sofronova 2003; Kaelke and Dawson 2005; Taiz and Zeiger 2006). Dean (2001) indicated that root allocation can be affected by stem competition because stems precede roots on the chain of carbohydrate sinks.
For future studies aimed at monitoring carbohydrate levels in trees, additional factors must be considered. For example, twigs exhibited less variation compared with other tissues analyzed and were easy to collect any time of year without causing considerable severe injury to the trees. Leaves are only available during the growing season and it requires being careful to obtain a homogenous sample during the dormant season. In the case of sampling roots and trunks, the tree sustains more damage that may disrupt physiological functions. In addition, fine root sampling can be complicated due to the partial loss of the sample during the washing process (Kaipiainen and Sofronova 2003). Changes depending on species have also been reported for carbohydrate partitioning (Newell et al. 2002). Based on the results of this study, trunk, root, twig, or leaf tissue, and can be used to monitor glucose and starch levels throughout the year; however, the seasonal variation in different parts of the tree and the different species should be taken into consideration.
Acknowledgments
We thank Environmental Design Inc. for supporting this research.
- Copyright © 2009, International Society of Arboriculture. All rights reserved.
LITERATURE CITED









{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.