
Abstract
The major flavonoids of the leaves of Gleditsia species are flavone C-glycosides, with sufficient interspecific differences in distribution to allow for the identification of certain species. G. triacanthos was readily distinguished from all other species by high concentrations of leucocyanidin and two catechins. The involvement of G. triacanthos in the parentage of G. x texana and the verification of an artificial hybrid of G. triacanthos were determined by chromatographic analyses and identification of the compounds peculiar to G. triacanthos.
Species of the genus Gleditsia are not widely cultivated in American arboreta or botanic gardens. On the other hand, thousands of thornless specimens of our native honeylocust (G. triacanthos L.) are planted every year in urban and suburban landscapes. These thornless trees may be grown from seed or produced by vegetative propagation from thornless trees or thornless portions of thorny trees. All such plants must be considered as G. triacanthos f. inermis Schneid1. Most of the thornless cultivars presently in the nursery trade have also been selected for fruitlessness, producing few if any functional female flowers.
The increase in popularity of the new cultivars of honeylocust over the past decade has been paralleled by an increase in the range of its most damaging insect pest, the mimosa webworm (Homadaula anisocentra Meyr.). Therefore, at the U.S. National Arboretum, a research project on Gleditsia was started in 1973 with the long-range goal of selecting or developing improved cultivars with a high degree of resistance to mimosa webworm. As a part of this project, biochemical studies were undertaken to determine the value of biochemical data in (1) identification or verification of species and hybrids, (2) determination of the nature of insect resistance.
The most recent monograph on Gleditsia (Gordon, 1966) recognizes 13 species and one natural hybrid, and the nomenclature of this work is followed here. Some of these species cannot be expected to be hardy in temperate zones of the United States, and many species are rare in cultivation. However, we have tried to collect and sample living material of all cultivated species, including a new species recently described from China.
Previous reports on Gleditsia leaf flavonoids have been rather limited. Bate-Smith (1962) did not detect the flavonols quercetin or kaempferol in acid-hydrolyzed leaf extracts of G. caspica Desf. However, he did report the presence of caffeic acid and a weak concentration of cyanidin (from leucocyanidin). Bate-Smith and Lerner (1954) found that G. triacanthos was strongly positive for cyanidin but that G. sinensis Lam. gave a negative reaction.
Materials and Methods
Sexually mature trees sampled in this survey were G. triacanthos, including the thornless cultivars ‘Imperial’ and ‘Moraine’ at the National Arboretum; G. japonica Miq. and G. x texana Sarg. from the Arnold Arboretum, Jamaica Plain, Massachusetts; G. aquatica Marsh, at the Morris Arboretum, Philadelphia, Pennsylvania; and a specimen identified as G. melanacantha Tang & Wang found growing at the USDA Plant Introduction Station in Glenn Dale, Maryland. In addition, a large, nonflowering specimen of G japonica at the National Arboretum was tested.
The remainder of the species, as well as all those mentioned above, were represented by one- or two-year-old seedlings. Seed was obtained from arboreta and botanic gardens throughout the world, either from their regular seed lists or through personal communication, Table 1 gives the primary sources. A total of 47 seedlots were tested, and in most cases we obtained seed of a given species from two or more sources. Sources other than those in the table included gardens in France, Germany, Portugal, and Tunisia.
Primary seed sources of Gleditsia species used in biochemical analyses.
Seed samples of all accessions were weighed, measured, color-coded, and stored dry in sealed containers in a refrigerator. Before sowing, the seed were soaked in concentrated sulfuric acid for one hour. Data on germination and seedling growth characteristics were recorded.
Seedling leaves for biochemical analyses were collected after the plants were three months old in the first growing season or during the second growing season. Leaf samples for flavonoid analyses, up to 30 grams fresh weight from mature trees and 5 to 10 grams for seedlings, were extracted in boiling 70% ethanol for one hour. Similar leaf samples were hydrolyzed in boiling 18% hydrochloric acid for 40 minutes to determine leucoanthocyanin content.
The ethanolic extracts were concentrated to about 1 ml extract for each gram of fresh leaf tissue. The extracts were allowed to cool and were cleared by centrifugation and stored in a refrigerator. For preliminary analyses, the extracts were spotted on Whatman No. 3MM filter paper (20 x 20 cm) and subjected to two-dimensional ascending chromatography in BAW (n-butanol:glacial acetic acid:water, 4:1:5, upper phase) and distilled water. These chromatograms, after being fumed with ammonia and viewed under ultraviolet light, provided tentative information on the number and behavior of compounds likely to be found in the extract. The various substances were purified by repeated banding and elution of one-dimensional runs in BAW, water, and 15% acetic acid, in whatever sequence was most appropriate for complete separation from other pigments.
Purified substances were then tested against known compounds by comparison and cochromatography in all three solvents and occasionally in TBA (tertiary butanol-glacial acetic acid-water, 3:1:1). Pure samples of vitexin, isovitexin, and iso-orientin were supplied by Dr. S. Asen, U.S. Department of Agriculture. Commercially available material of d-catechin, epicatechin, chlorogenic acid, caffeic acid, and several other compounds were also used in this manner.
Detection and characterization of spots was sometimes aided by such chromogenic sprays as diazotized p-nitroaniline and tetrazotized benzidine.
The Rf values of purified C-glycoflavones were not altered by acid hydrolysis, and no sugars were detected following such treatment. C-glycoflavones have been reported as unusually resistant to acid hydrolysis.
In addition to Rf values and cochromatography, great reliance was placed on the ultraviolet absorption spectra-in 95% ethanol (compared with data of Harborne, 1967) or methanol (compared with data of Mabry et al. 1970). Spectral shifts with a variety of additives (sodium acetate, boric acid, aluminum chloride) also aided in identification.
The presence of leucoanthocyanins in acid-hydrolyzed leaf samples could sometimes be judged by the red coloration of the extract, especially when leucoanthocyanin content was high. The raw extract was first extracted with isoamyl alcohol, and this layer was washed twice with 1 % HCL. The addition of a five-fold volume of benzene formed a one-phase system. After the addition of more 1 % HCL the red pigment appeared in the lower aqueous layer. The purified pigment was recovered from this layer with isoamyl alcohol, and the anthocyanidins were identified by paper chromatography in Forestal solvent (acetic acid:HCL:water, 30:3:10) and by their absorption spectra.
Results and Discussion
Figure 1 shows composite chromatogram of compounds found in the various Gleditsia species. Spots other than flavonoids are included for the sake of completeness and for the use of future investigations. Table 2 shows the Gleditsia species we have studied, with notes on the presence or absence of the various substances.
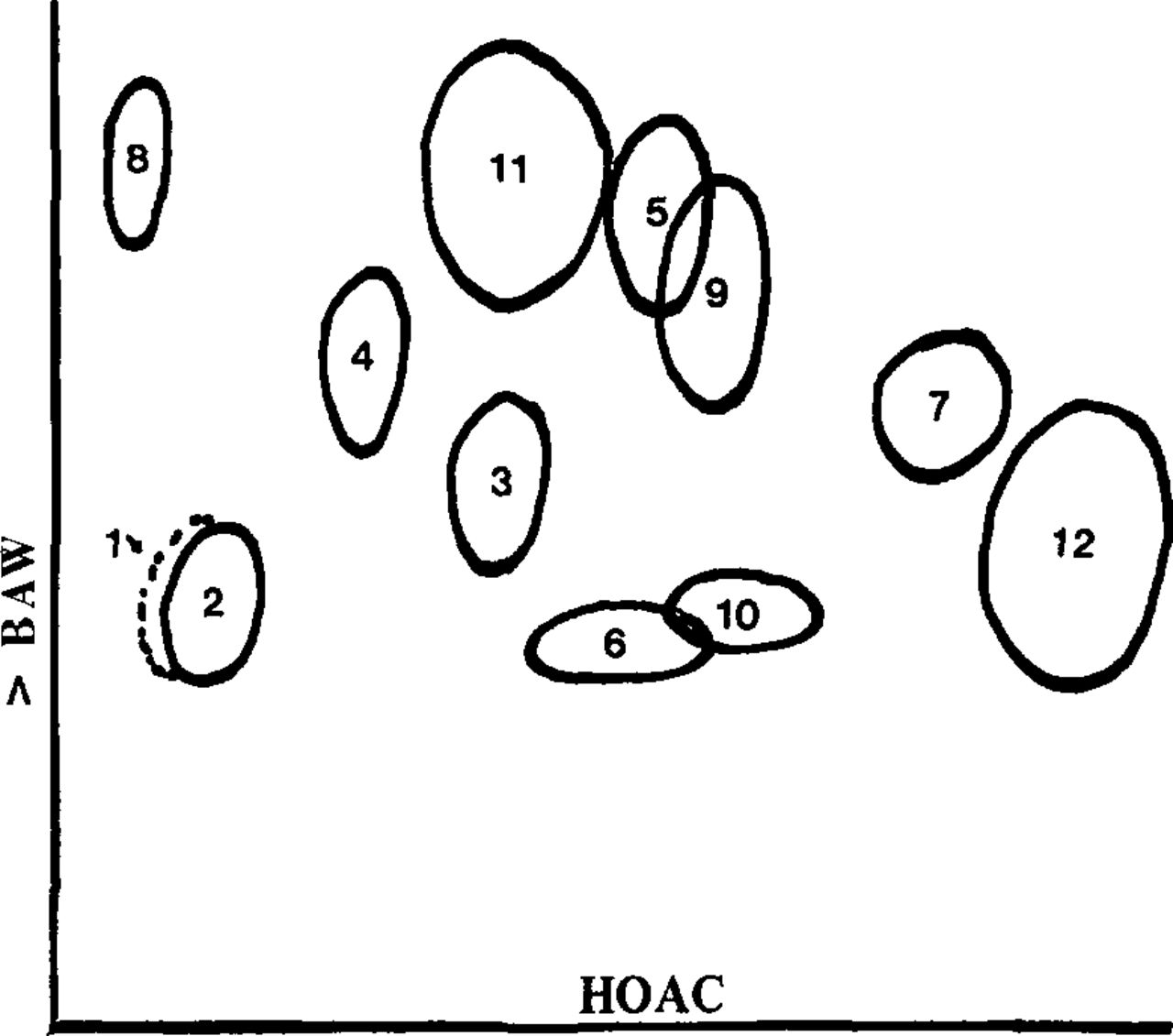
Composite two-dimensional chromatogram of Gleditsia leaf flavonoids and related substances. Numbered substances are discussed in the text and their distribution is given in Table 2.
Flavonoids and related compounds found in Gleditsia leaves.
The characteristics and identification of the various substances are discussed below. Some substances (#3, 4, 5, 9, 11 and 12) were compared with authentic materials, and such identifications were highly probable. Little additional data are given on them, because reference works contain full details. For other substances, some pertinent information is given.
Cyanidin (CY) is not pictured on the chromatogram because of the different solvent system used in its identification. It is assumed that this compound resulted largely from the hydrolysis of condensed tannins in the leaves.
The following substances were identified:
Substance #1. A substance that fluoresces yellow and has similar Rf values to orientin. Not studied further.
Substance #2. Probably orientin (luteolin 8-C-glucoside). Formerly called lutexin.
Substance #3. Iso-orientin (luteolin 6-C-glucoside). Formerly called lutonaretin.
Substance #4. Vitexin (apigenin 8-C-glucoside).
Substance #5. Isovitexin (apigenin 6-C-glucoside). Formerly called saponaretin.
Substance #6. Probably a vitexin derivative, and possibly a C-diglycoside. Rf values of 0.32 (BAW) and 0.48 (HOAC) are inconclusive, because Harborne (1967) and Mabry et al (1970) present very different values.
Substance #7. Probably rhamnosylvitexin (apigenin 8-C rhamnoglucoside). Rf values of 0.52 (BAW) and 0.74 (HOAC). UV spectral shifts similar to those reported by Mabry et al. (1970).
Substance #8. Possibly luteolin as a free flavone. Not studied further.
Substance #9. d-Catechin. Rf values 0.60 (BAW), 0.54 (HOAC), 0.33 (H2O). Identity confirmed by co-chromatography with pure compounds. UV absorbtion maximum at 278 nm.
Substance #10. Another catechin-type compound. Does not appear to be any of the common tea components. Rf values 0.35 (BAW), 0.58 (HOAC), 0.48 (H2O).
Substance #11. Caffeic acid, probably trans isomer. Rf values 0.70 (BAW), 0.37 (HOAC), Blue fluorenscent in UV.
Substance #12. Chlorogenic acid. Gives green-yellow fluorescence under UV after fuming with ammonia vapor. Rf values 0.41 (BAW), 0.86 (HOAC). Co-chromatographed with commercial chlorogenic acid.
The major flavonoids of the genus Gleditsia, as in some other legumes, are glycoflavones, in which the sugar is attached by a carbon-carbon bond to the flavone nucleus. Species differences were not marked and no signigicant species-specific glycoflavone compounds were found, although thorough analyses and comparisons were used to aid in species identification on material of doubtful origin.
However, from a practical viewpoint, other flavonoids, such as catechins and leucoan-thocyanidin, were used in distinguishing G. triacanthos from all other species studied. Flavonoid tannins are polymeric condensation products of catechins (flavan-3-ols) and leucoan-thocyanins (flavan-3:4-diols) and they are usually accompanied in the plant by their monomeric precursors. Thus, although the monomeric catechins could be easily determined in the alcoholic leaf extracts, the leucocyanidin could best be identified after tannin cleavage by acid hydrolysis.
G. x texana has long been considered a natural hybrid between G. aquatica and G. triacanthos. The chemical evidence from mature trees and open-pollinated seedlings of G. x texana indicated that G. triacanthos was definitely involved in the parentage of that species.
When leaf tissue from G. triacanthos was hydrolyzed, the solution turned red after about 10 minutes of boiling. The degree of “redness” was less for mature trees of G. x texana and still less for seedlings derived from G. texana. Although no precise quantitative data were taken, the crude extracts and the purified material clearly showed that the amount of leucocyanidin diminished with the reduction in the contribution of G. triacanthos germplasm.
No reddish color was apparent in the acid-hydrolyzed extract of G. japonica, and leucocyanidin could only be determined after purification.
Leaves of the Chinese G. melanacantha were similar to G. triacanthos in giving a marked red coloration to the solvent during acid hydrolysis. After purification, leucocyanidin was identified chromatographically. In addition, what appeared to be two or three other leucoanthocyanidins were detected, although all had absorbtion peaks similar to cyanidin. No catechin compounds were found in G. melanacantha, and it was possible that the leucoanthocyanins in this species were derived from monomeric flavan-3:4 diols.
A few young hybrid seedlings of G. melanacantha x G. triacanthos derived from controlled pollination were also studied. True hybridity of these plants was verified by the presence of the catechin compounds in the alcoholic extract.
It may be noteworthy that we also found these catechin compounds in the leaves of Albizia julibrizzin Durazz. and A. kalkora Prain, two other hosts of the mimosa webworm. Apparently the high tannin content in the leaves of both honeylocust and “mimosa” is no deterrent to feeding by this insect pest.
Conclusions
In any breeding program to develop superior and insect-resistant cultivars of honeylocust, our native species G. triacanthos should be the key species. The chemical characteristics peculiar to G. triacanthos should allow rapid chromatographic verification of interspecific hybrids involving G. triacanthos as the male parent. The fact that most selected thornless cultivars of G. triacanthos are also predominantly male-flowering virtually eliminates the possibility of reciprocal crosses and suggests that this species will be more often used as a male. In view of the marked morphological similarity among many Gleditsia species, chemical criteria for judging hybridity will be critical factors in progeny evaluation.
Footnotes
↵1 Botanical authority according to Isely (1975).
- © 1977, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.