
Abstract
Numerous studies have shown that ectomycorrhizal fungi can profoundly affect conifer performance by facilitating nutrient and water uptake, maintaining soil structure, and protecting roots from pathogens and environmental extremes. However, fertilizing and irrigating practices in seedling production nurseries are very different than field conditions at harsh outplanting sites. More information is needed on the ability of specific mycorrhizal fungi to establish at the nursery and improve seedling performance in the outplanted environment. This study was conducted to test the ability of a specific ectomycorrhizal fungus, Rhizopogon rubescens, inoculated onto the root systems of plug-1 ponderosa pine (Pinus ponderosa) seedlings grown in fumigated and nonfumigated bare-root nursery beds to influence conifer establishment on two harsh, dry sites in southwest Oregon, U.S. After outplanting, survival of Rhizopogon-inoculated seedlings were significantly higher than noninoculated seedlings at both field sites (p < 0.05). Survival averaged 93% for Rhizopogon-inoculated seedlings and 37% for noninoculated seedlings at the Central Point site. Survival averaged 71% for Rhizopogon-inoculated seedlings and 41% for noninoculated seedlings at the Applegate site. Field survival did not differ significantly for ponderosa pine seedlings grown in fumigated compared to nonfumigated beds. Seedling height did not differ significantly between Rhizopogon-inoculated and noninoculated ponderosa pine seedlings or fumigated and nonfumigated beds in the nursery or outplanting sites. Foliar analysis at the Applegate site indicated significantly higher phosphorous contents for Rhizopogon-inoculated seedlings. Results from this study indicate that Rhizopogon inoculated plug-1 ponderosa pine survive at a much higher rate on dry, harsh sites in southwest Oregon. Poor survival by noninoculated pine seedlings grown in both fumigated and nonfumigated beds and outplanted on harsh sites indicate that field survival should be considered one of the more important criteria for selection of Rhizopogon species suitable for nursery inoculation.
- Rhizopogon spp.
- ponderosa pine
- Pinus ponderosa; mycorrhizae
- mycorrhizal fungi
- spore inoculation
- fumigation
- transplanting
- survival
- drought tolerance
- conifer nursery
Throughout the western United States, ponderosa pine (Pinus ponderosa Dougl. ex Laws.) trees are planted extensively on a variety of sites in urban, suburban, and forest environments. Disturbed, compacted soils and hot, dry sites are commonly encountered. First-year mortality of planted trees can be high under harsh conditions (Preest 1977; Peterson and Newton 1985; Amaranthus and Perry 1987; Amaranthus and Malajczuk 2001) and foresters, landscapers, and arborists are always interested in cultural practices at the nursery that may improve tree survival and performance. Conifer tree establishment depends on rapid root and ectomycorrhizal formation on dry sites difficult to regenerate (Amaranthus and Perry 1989). In the arid western United States, transpiration potential during the growing season can exceed soil water availability, killing or reducing growth of nonirrigated seedlings. Ectomycorrhizae enhance water uptake by their hosts (Trappe and Fogel 1977; Reid 1979; Parke et al. 1983), although tolerance to low water potentials vary widely among mycorrhizal species (Mexal and Reid 1973; Theodorou 1978; Parke et al. 1983). Amaranthus and Malajczuk (2001) found Rhizopogon rubescens mycorrhizal colonization of longleaf pine seedlings (Pinus palustris Mill.) significantly reduced plant moisture stress at low soil moisture levels. Plant moisture stress levels averaged 78% higher for noninoculated seedlings compared to Rhizopogon-inoculated seedlings at low soil moistures. Theodorou and Bowen (1970) also observed that P. radiata seedlings inoculated with R. luteolus survived a particularly dry summer better than nontreated seedlings. Sands and Theodorou (1978) found leaf water potentials of Rhizopogon-inoculated seedlings were lower than for noninoculated seedlings.
Numerous studies have shown improvement of outplanting performance of Rhizopogon-inoculated seedlings in conifer establishment. (Volkart 1964; Theodorou and Bowen 1970; Theodorou 1971; Momoh 1976; Castellano and Trappe 1985; Ekwebelam and Odeyinde 1985, Amaranthus and Perry 1989, Castellano 1996). Nutrient acquisition is considered a major factor improving seedling growth. Significantly increased uptake of phosphorus has been reported for mycorrhizal inoculated conifer seedlings (Theodorou and Bowen 1970; Lamb and Richards 1971, 1974; Skinner and Bowen 1974a, 1974b; Chu-Chou 1979, Chu-Chou and Grace 1985), as well as potassium (Theodorou and Bowen 1970; Lamb and Richards 1971), sodium (Melin et al. 1958), total nitrogen (Chu-Chou and Grace 1985), and ammonia forms of N (Finlay et al. 1988).
Considerable effort and expense is directed toward site preparation at many suburban and urban sites. Mycorrhizal inoculum density and viability are often reduced as of a result of site preparation activities (Amaranthus et al. 1994, 1996; Dumroese et al. 1998). Amaranthus et al. (1996) found significant reductions in mycorrhizal abundance and diversity, including Rhizopogon spp. with moderate to high levels of organic-matter removal and soil compaction. Rhizopogon spp. produce belowground fruiting bodies that require animals to spread spores via fecal pellets. They do not produce airborne spores, which makes it unlikely that Rhizopogon mycorrhizal fungi would quickly be introduced from surrounding natural areas to disturbed urban and suburban sites.
The role of mycorrhizal fungi in the health and vigor of trees in stressful environments is well documented. However, more information is needed regarding establishing specific native mycorrhizal fungi in conifer tree nursery environments to increase seedling survival on harsh planting sites. Nursery inoculation of mycorrhizal fungi selected to promote survival and growth in a dry forest, suburban, or urban environment could be an important tool for foresters, landscapers, and arborists. This study was conducted to test the ability of a specific ectomycorrhizal fungus, R. rubescens, inoculated onto the root systems of plug-1 ponderosa pine seedlings grown in fumigated and nonfumigated bare-root nursery beds to influence outplanting performance on two harsh dry sites in southwest Oregon.
MATERIALS AND METHODS
Nursery Procedures
On July 1, 1999, ponderosa pine seeds were sown in 2 in3 cells in Stryoblock™ containers at the J. Herbert Stone Nursery in Central Point, Oregon. On July 12, emerging ponderosa pine seedlings were inoculated with 100,000 spores each of the mycorrhizal fungus R. rubescens using an injection of a liquid suspension via a traveling irrigation boom. Spores were applied as a soil drench following maceration of R. rubescens sporocarps for 10 minutes in distilled water. Spore concentrations were determined with a haemacytometer. Foliar fertilizer (250 ppm N, 31 ppm P, and 158 ppm K plus micronutrients) was applied each irrigation during the rapid growth phase. Greenhouse temperatures were held between 65°F and 75°F. In early September 1999, seedlings were hardened-off by reducing irrigation and changing the fertilizer rates (50 ppm N, 60 ppm P, and 150 ppm K plus micronutrients). On September 22, 1999, ponderosa pine seedlings were inoculated again with 100,000 spores each of R. rubescens using the same inoculation procedure. No pesticides were used on the crop during this period.
On September 29, 1999, Rhizopogon-inoculated and noninoculated ponderosa pine were extracted and transplanted into fumigated and nonfumigated bare-root nursery beds at J. Herbert Stone Nursery. At that time 6 Rhizopogon-inoculated and noninoculated container seedlings were examined for the percentage of colonization by the mycorrhizal fungus R. rubescens. Mean colonization by R. rubescens on inoculated seedlings was 8%, while no Rhizopogon was present on noninoculated seedlings. Fumigated beds were treated with methyl bromide the prior year, while nonfumigated beds had not been fumigated since October 1996. Prior to transplanting, 228 kg per hectare of ammonium phosphate (16-20-0) and 228 kg per hectare of potassium sulfate (0-0-50-53) were incorporated into the soils.
After transplanting, seedlings were grown using standard cultural practices for bare-root production. Seedlings were fertilized with 197 kg of N in the form of ammonium nitrate and ammonium sulfate during spring 2000. Root wrenching occurred four times during spring and summer 2000. No pesticides were used on the transplant crop. Seedlings were lifted on January 8, 2001, and those not meeting minimum seedling diameters of 5 mm and seedling height of 13 cm were discarded. Seedlings with poorly developed root systems or J-roots were also removed. Diameters, heights, and root volumes of seedlings to be outplanted were measured on 30 seedlings each from fumigated, nonfumigated, Rhizopogon-inoculated, and noninoculated plots. No significant differences in seedling diameters, heights, or root volumes (p = 0.05) were measured between treatments. Seedlings were placed in cold storage for 4 months until they were outplanted.
Outplanting Procedures
Seedlings were outplanted in two locations in southwest Oregon—the Central Point and Applegate study sites. The Applegate study site is in a small valley at 385 m elevation in the Siskiyou Mountains. Historical annual precipitation averages 650 mm, less than 10% of which falls from mid-May through mid-September. Soils are fine loamy, mixed mesic Ultic Haploxeralfs, 60 to 100 cm deep, formed in granitic colluvium and underlain by weathered granitic bedrock. Soils are classified in the Holland series. Surface layers (to 18 cm) are dark grayish brown to brown sandy loams. Percentages of sand, silt, and clay are 52, 24, and 24 respectively. The study area is on a southwest-facing, gentle (< 5%) toe slope just above the valley bottom. Soil moisture was at field capacity (28%) at the time of outplanting ponderosa pine.
The Central Point site is located at the J. Herbert Stone Nursery near Central Point, Oregon, at 426 m elevation on a nearly level slope (< 5%). Historical annual precipitation averages 500 mm, less than 10% of which falls from mid-May through mid-September. Soils are coarse-loamy, mixed, mesic Pachic Haploxerolls, more than 100 cm deep, formed from granitic and metamorphic alluvium. They are classified in the Central Point series. Surface layers are black, sandy loams about 40 cm thick. The planting site is located in an unirrigated field. Soil moisture at the time of planting was at field capacity (15%) at time of planting.
Seedlings were outplanted on May 9, 2001, at both sites. At each site, 16 plots (2 × 2 m) were established for field assessment of Rhizopogon-inoculated/noninoculated and fumigated/nonfumigated treatments. Each area was planted with randomly assigned treatments of nine seedlings each in a 3 × 3 array at 40 cm spacing. Each of the 16 plots were separated by 1 m buffers. The treatments were (1) Rhizopogon-inoculated/fumigated beds, (2) noninoculated/ fumigated beds; (3) Rhizopogon-inoculated/nonfumigated beds; and (4) noninoculated/nonfumigated beds. Each treatment was replicated four times at each site. In September 2001, tree heights at ground line were measured on surviving seedlings and the number of surviving seedlings tallied for each treatment area.
Laboratory Procedures
Before outplanting, five randomly selected seedlings were examined for presence of Rhizopogon ectomycorrhizae for each of the 16 plots. Roots were gently washed free of soil and extraneous material and subsampled in three cross sections, 1.5 cm broad, of the entire root systems in upper, middle and lower positions, respectively. All active tips were tallied as Rhizopogon, other mycorrhizal or nonmycorrhizal from characteristics observed through a dissecting microscope (2× by 10× magnification). Mycorrhizal tips were separated by type according to characteristics observable through a dissecting microscope (2× to 10× magnification). Rhizopogon mycorrhizae identification was verified using color, surface appearance, branching, morphology, degree of swelling, length, and characteristics of rhizomorphs. Rhizopogon rubescens mycorrhizae were creamy white and developed a gradient of yellow and reddish coloration with maturity and upon bruising. The R. rubescens mycorrhizae had a two-layered mantle and abundant rhizomorphs developing a compact coralloid morphology with maturity. In September 2001, pine needle samples were collected from four randomly selected seedlings from each treatment at the Applegate site. Samples were analyzed for total N, P, and K (Kjeldahl digest with ammonia and orthophosphate read on an autoanalyzer).
Statistical Procedures
The experimental design was a randomized block. ANOVA was selected as the primary analysis technique. ANOVAs were performed separately for seedling survival, height, foliar nutrients, and mycorrhizal colonization (Steel and Torrie 1980). Means comparisons were calculated using Fisher’s LSD. Residuals from the performed ANOVAs were examined using, normal probability plots, tests that the residuals come from normal distributions, and plots of residuals versus predicted values. Before analysis, data were logarithmically transformed to compensate for log-normally distributed values (Steel and Torrie 1980).
RESULTS AND DISCUSSION
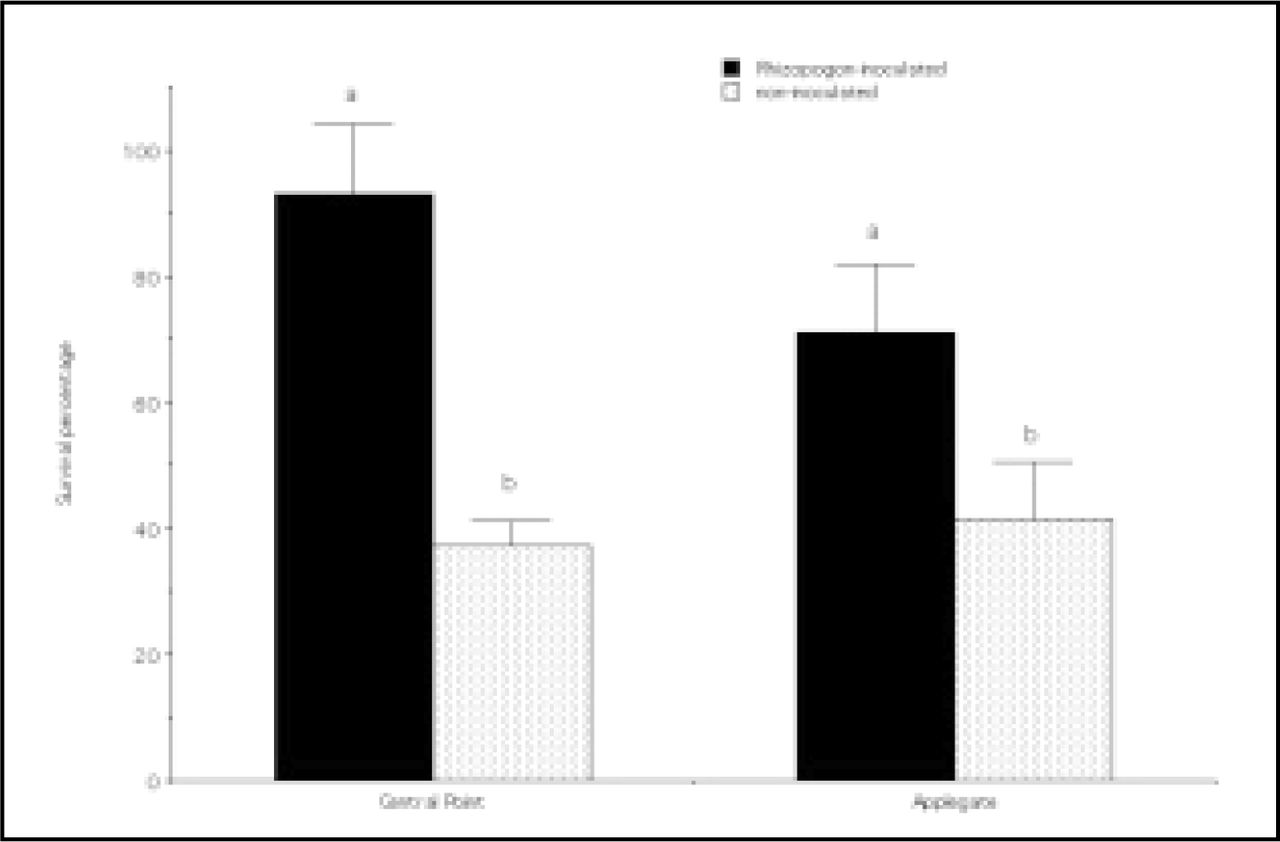
Ponderosa pine outplanting survival following Rhizopogon inoculation was significantly higher compared to noninoculated seedlings (p < 0.05; Figure 1 and Figure 2*). The average seedling survival for Rhizopogon-inoculated seedlings was 93% compared to 37% for noninoculated seedlings for the Central Point site. The average seedling survival for Rhizopogon-inoculated seedlings was 71% compared to 41% for noninoculated seedlings for the Applegate site. Seedling height at the time of outplanting and after the first growing season in the outplanting environment was not significantly different for any treatment and site combination (Figure 3).

Seedling survival after outplanting Rhizopogon-inoculated and noninoculated ponderosa pine seedlings from the outplanting sites at the USDA Forest Service J. Herbert Stone Nursery at Central Point and Applegate, Oregon. Bars represent means, and vertical lines represent standard errors. Means that share the same letter do not differ by Fishers LSD test, P = 0.05.

Photograph of one Rhizopogon-inoculated plot (top) and noninoculated plot (bottom) at the Central Point site.

Seedling height after outplanting Rhizopogon-inoculated and noninoculated ponderosa pine seedlings from fumigated and nonfumigated outplanting sites at the USDA Forest Service J. Herbert Stone Nursery at Central Point and Applegate, Oregon. Bars represent means, and vertical lines represent standard errors. Means that share the same letter do not differ by Fishers LSD test, P = 0.05.
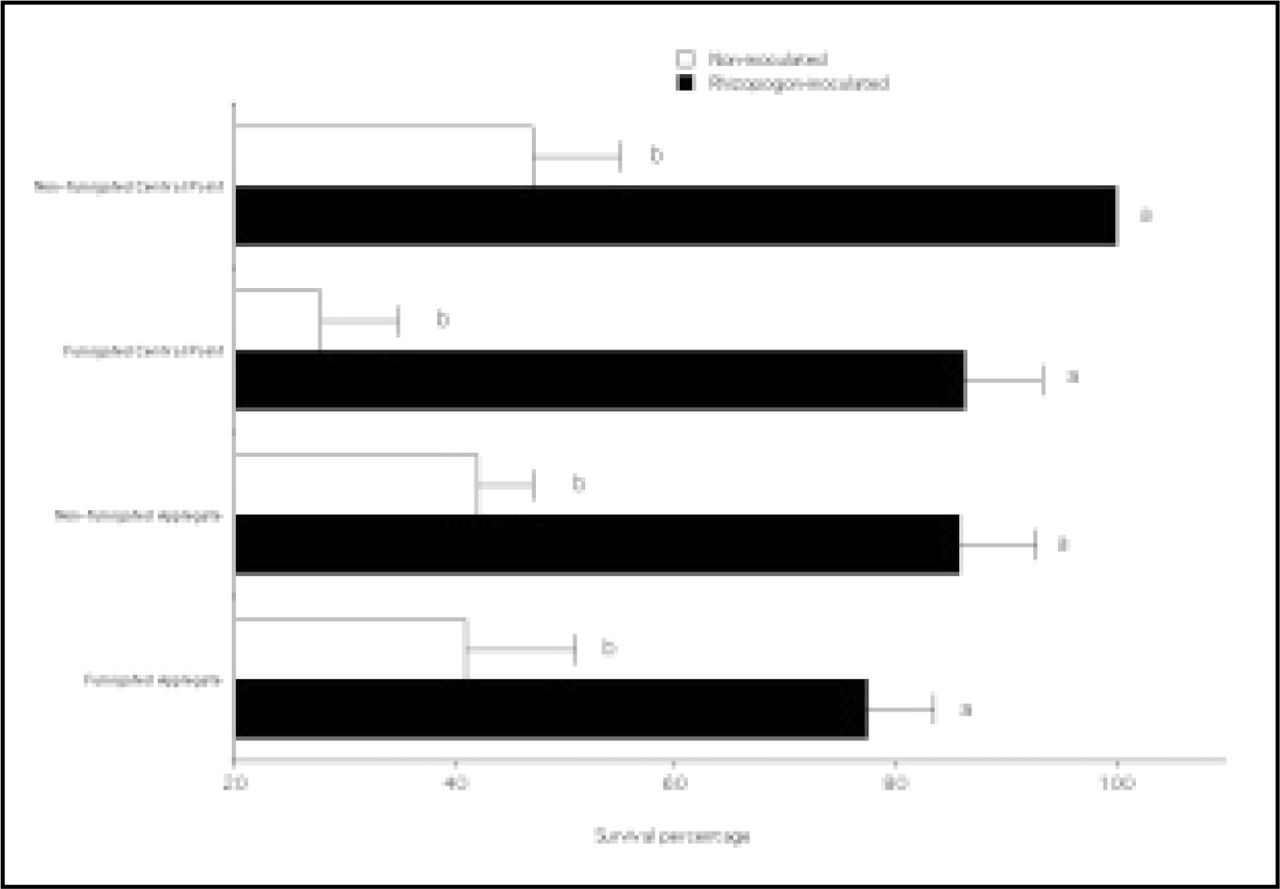
Rhizopogon mycorrhizal colonization was significantly higher on Rhizopogon-inoculated seedlings compared to noninoculated seedlings (Figure 4). Seedlings from fumigated beds had higher Rhizopogon colonization (28%) compared to nonfumigated beds (17%) and noninoculated seedlings from fumigated and nonfumigated beds (1%). However, there were no statistical differences between survival of Rhizopogon-inoculated seedlings from fumigated and nonfumigated beds (Figure 5).

Rhizopogon percentage mycorrhizal colonization before outplanting Rhizopogon-inoculated and noninoculated ponderosa pine seedlings from fumigated and nonfumigated outplanting sites at the USDA Forest Service J Herbert Stone Nursery at Central Point and Applegate, Oregon. Bars represent means, and vertical lines represent standard errors. Means that share the same letter do not differ by Fishers LSD test, P = 0.05.

Survival percentage after outplanting Rhizopogon-inoculated and noninoculated ponderosa pine seedlings from fumigated and nonfumigated outplanting sites at the USDA Forest Service J Herbert Stone Nursery at Central Point and Applegate, Oregon. Bars represent means, and vertical lines represent standard errors. Means that share the same letter do not differ by Fishers LSD test, P = 0.05.
Foliar phosphorous contents were significantly higher on Rhizopogon-inoculated seedlings outplanted from both fumigated beds and nonfumigated beds (Figure 6). Phosphorous percentages were 90% and 60% higher on Rhizopogon-inoculated seedlings outplanted from fumigated beds and nonfumigated beds, respectively, compared to noninoculated seedlings. Foliar nitrogen and potassium levels were higher but not significantly different from noninoculated seedlings (p < 0.05) from fumigated and nonfumigated beds planted at the Applegate site.

Foliar NPK percentage after outplanting Rhizopogon-inoculated and noninoculated ponderosa pine seedlings from fumigated and nonfumigated outplanting sites at the USDA Forest Service J Herbert Stone Nursery at Central Point and Applegate, Oregon. Bars represent means, and vertical lines represent standard errors. Means that share the same letter do not differ by Fishers LSD test, P = 0.05.
Seedlings planted in the western United States are usually subjected to low rainfalls and high temperatures during the summer months after spring outplanting. In our study, both outplanting areas represent typical harsh sites encountered in southwestern Oregon. Rainfall in the months following outplanting of our study was very low (56 mm between May 1 and October 1, 2001), and afternoon ambient air temperatures were high (temperatures exceeded 32°C on 49 days between May 1 and October 1, 2001). As a result, many seedlings at the Central Point site showed signs of wilting and mortality as early as the end of June, 6 weeks after outplanting. Seedlings continued to die as the summer progressed and soils dried out. At both outplanting sites, ponderosa pine seedlings with roots colonized by Rhizopogon mycorrhizae, however, survived significantly better than noncolonized seedlings. This finding may be related to the properties and functions of Rhizopogon that decrease plant moisture stress and promote drought tolerance as soils dry out. Amaranthus and Malaljczuk (2001) found that at high soil moisture contents, there were no significant differences in plant moisture stress between Rhizopogon-inoculated and noninoculated of longleaf pine seedlings. But as soils dried down to as low as 4% soil moisture, differences in plant moisture stress between inoculated and noninoculated seedlings became significant, with inoculated seedlings averaging 9.8 bars and noninoculated seedlings averaging 20 bars.
The mechanism by which Rhizopogon mycorrhizae reduce plant moisture stress in dry soil conditions is becoming better understood. On examination of the excavated Rhizopogon-inoculated pine seedling root systems, we observed spongy mycorrhizal mantles and abundant rhizomorphs (Figure 7). Hydration and slow release of water to the tree from spongy fungus mantles and rhizomorphs during drought conditions could buffer seedlings and help reduce plant moisture stress. Spongy mantles and rhizomorphs have been noted and described in numerous Rhizopogon studies (Massicotte et al. 1994; Molina and Trappe 1994; Agerer et al. 1996). Rhizomorphs play an important role in water storage and movement (Duddridge et al. 1980; Brownlee et al. 1983; Read and Boyd 1986). Parke et al. (1983) and Dosskey et al. (1990) demonstrated enhanced tolerance to drought stress of Douglas-fir (Pseudotsuga menziesii) seedlings inoculated with R. vinicolor and attribute this enhancement in part to rhizomorph production and function in water storage and transport.

Photograph of Rhizopogon-mycorrhizae with spongy mantle and abundant rhizomorphs.
Benefits of inoculating seedlings with Rhizopogon mycorrhizae might not be apparent to managers of bareroot and container nurseries who are trying to produce larger seedlings. The lack of aboveground differences between inoculated and noninoculated seedlings in this study at the nursery is commonly observed by many nursery managers who inoculate with ectomycorrihzae. Why aboveground differences are not apparent in nurseries could be due to the relatively low moisture stress and high soil nutrient levels typical of nursery environments. In our study, nursery seedlings were never subjected to stresses that exceeded a pre-dawn moisture stress of 10 bars. Soils were kept moist for most of the time that seedlings were in bare-root beds except for a 4 to 6 week period in late summer 2000 when the soils were allowed to dry to induce seedling hardening. Since mycorrhizae support seedlings when moisture and nutrients are limiting, their function in the nursery environment might be of limited advantage.
Outplanting benefits of nursery Rhizopogon-inoculation, however, are well documented. Results from studies throughout the world have demonstrated the importance of Rhizopogon spp. as ectomycorrhizal symbionts in the successful establishment of conifers. As early as 1927, Kessel recognized R. luteolus as being among the first fungi to fruit in association with scattered “healthy” radiata pine in Australian nurseries. Chu-Chou (1979) reemphasized the importance of Rhizopogon in conifer plantations and nurseries in New Zealand. Chu-Chou and Grace (1981, 1983) later discovered R. vinicolor and R. parksii to be dominant ectomycorrhizal fungi of introduced Douglas-fir seedlings in nurseries and plantations. In Nigeria, Momoh (1976) has found R. luteolus associated with vigorously growing introduced pines. Rhizopogon ectomycorrhizae also have been associated with the establishment of conifers in Africa (Donald 1975; Fogel 1980; Ivory 1980), Puerto Rico (Volkart 1964), Europe (Levisohn 1956, 1965; Gross et al. 1980; Jansen and de Vries 1989; Parlade and Alvarez 1993; Parlade et al. 1996), New Zealand (Birch 1937; Chu-Chou and Grace 1981, 1983), South America (Garrido 1986), and the United States (Baxter 1928). More recently, the importance of Rhizopogon in increased seedling performance in the field following nursery inoculation was demonstrated in the Oregon Coast Range. Amaranthus and Perry (1994) inoculated nursery-grown containerized Douglas-fir seedlings from six families with spores of R. vinicolor. Inoculated and noninoculated seedlings from all families were outplanted in the Oregon Coast Range. Rhizopogon vinicolor-colonized seedlings from all families had significantly greater height growth (six of six families) and basal area growth (five of six families) compared to noninoculated seedlings.
The finding of no significant difference between survival of fumigated and nonfumigated seedlings has implications to nurseries that are moving away from using soil sterilants. Soil fumigation with broad-spectrum biocides is a nonselective means of killing soilborne pathogens in tree seedling nurseries (Linderman 1994; Marx et al. 1979). Nursery practices that utilize methyl bromide or other soil sterilants are known to reduce or eliminate mycorrhizal fungi (Lee and Koo 1985; Davies 2002). At J. Herbert Stone Nursery, the soils of the nonfumigated treatments had been fumigated 3 years before the seedlings were transplanted in this study. During this period, changes in soil biological composition and the reintroduction of mycorrhizae, including Rhizopogon spp., have been slow. Rhizopogon spp. produce belowground fruiting bodies that do not disperse their spores through the air, thus making reintroduction from natural areas more difficult. The results of our study suggest that if mycorrhizae-colonized seedlings are to be produced in bare-root fields, inoculation with specific mycorrhizae will be necessary until a desirable population of mycorrhizae becomes established in nonfumigated nursery fields.
The importance of ectomycorrhizal fungi in the uptake and translocation of nutrients to their host plants has been the underlying principal in numerous studies of conifers. Of particular importance is the role of ectomycorrhizal fungi in phosphorous nutrition. Ectomycorrhizal fungi produce acid phosphatases, a special type of root exudate that hydrolyses organically bound phosphorous. Bowen and Theodorou (1968) found that R. roseolus cultures were able to solubilize rock phosphate, and Theodorou (1968) also indirectly showed substantial phosphatase activity by R. roseolus. Skinner and Bowen (1974) demonstrated the uptake and translocation of phosphate via mycelial strands of pine mycorrhizae. Ho and Trappe (1987) found that six Rhizopogon spp. tested produced acid and alkaline phosphatases as well as nitrate reductase, an enzyme that aids the acquisition of nitrogen. Ho and Trappe (1980) report that R. vinicolor produced high levels of nitrate reductase compared to other ectomycorrhizal fungi. In our study, Rhizopogon-inoculated pine seedlings had significantly increased levels of foliar phosphorous compared to noninoculated seedlings in one growing season after outplanting. Rhizopogon-inoculated seedlings also had increased foliar levels of nitrogen and potassium, but differences were not significantly different. The best-documented mycorrhizal effect in the literature is that mycorrhizal plants take up more soil phosphorous than nonmycorrhizal plants do. In our study, we see a similar P effect, but it is unlikely that improved phosphorous nutrition had a substantial effect on seedling survival.
Mycorrhizal fungi play a key role in the health and vigor of trees in stressful environments. In southwest Oregon, dry spring and summer conditions often result in significant conifer mortality upon outplanting. Results from this study indicate that Rhizopogon inoculation at the nursery can help seedlings survive and establish on difficult sites. Nursery inoculation of specific mycorrhizal fungi, such as Rhizopogon spp., selected to promote survival and growth in dry and disturbed forest, suburban, and urban environments could be an important tool for foresters, landscapers, and arborists.
Acknowledgments
We are grateful to Roy Young, Dawn Huck, Steve Feigner, David Davis, and Johan Visser for invaluable assistance in the field. The authors thank to Drs. Tom Landis and Jim Trappe for their comments on drafts of the manuscript.
Footnotes
↵* Figures in this article are shown on pages 204–207.
- © 2003, International Society of Arboriculture. All rights reserved.
LITERATURE CITED
In this issue









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.