
Abstract
Systemic acquired resistance (SAR) is the phenomenon whereby a plant’s own defense mechanisms are induced by prior treatment with either a biological or chemical agent. The concept of SAR has been widely recognized and studied for the past 100 years in relation to increasing resistance to fungal, bacterial, and viral pathogens of economically important crop plants. The use of SAR as part of a disease management strategy in relation to urban forestry has received little scientific investigation despite offering some potential for tree pathogen control. The objectives of this paper are to discuss the concepts of SAR and, where applicable, indicate how SAR may be incorporated into a disease management system by urban foresters. Areas discussed include historical background, persistence of SAR, mechanisms of SAR, induction of SAR by biological organisms and chemicals, interplanting with flowering woody plants to promote an SAR response, and SAR in woody plants.
- Induced resistance
- integrated disease management
- disease control
- plant-pathogen interactions
- fungicides
- systemic acquired resistance
- SAR
- salicylic acid
- isonicotinic acid
- plant activator
During their life cycle, urban trees are susceptible to a broad range of pathogenic fungi which, if uncontrolled, can result in high mortality rates. Infection can be via the roots (e.g., Phytophthora cinnamomi and Armillaria mellea, which interfere with the absorption of water and nutrients from the soil), via leaves (e.g., leaf spots [Gnomonia platani, G. veneta], which infect foliage and reduce photosynthesis), via flowers (e.g., fungal blights that attack the flowers and affect reproduction), and via fruit rots such as Monilinia fructicola, which interfere with reproduction and/ or storage of food reserves. Indeed, tree diseases can be so problematic as to limit the type of tree species that can grow in a large geographic area. The American chestnut (Castanea dentata) was annihilated in North America as a timber tree by chestnut blight (Dierauf et al. 1997), and the American elm is being eliminated as a shade tree by Dutch elm disease (Stipes and Campana 1981).
The problem of tree diseases can be further exacerbated because trees are often already weakened by the wide range of abiotic stresses they encounter within a town or city environment (e.g., root hypoxia, salinity, and drought). Trees in this weakened state are more susceptible to a range of pathogens that under normal circumstances do not pose a threat to their biology. For example, birch trees planted in the south of England tend to suffer from drought stress on an annual basis. This in turn makes them more susceptible to rust pathogens (Cronartium spp.). Such a problem is rarely encountered in Scotland where cooler, milder growing conditions prevail.
Presently, foresters tend to control these diseases primarily through the use of chemicals or by pruning diseased tissue and burning the arisings (British Standards Institute 1989). Tolerance to fungicides in, for example, the brown rot fungus or Monilinia fructicola (Agrios 1997); failure of fungicides to control diseases once a tree is infected, for example, by Armillaria mellea; increased legislative restrictions of the use of agrochemicals; and failure to achieve even distribution of chemicals throughout a tree canopy means that new techniques of pathogen control are required.
It is widely known that plants can defend themselves against pathogen infection through a variety of mechanisms that can be either local, constitutive, or inducible (Franceschi et al. 1998, 2000). Inducible resistance mechanisms such as systemic acquired resistance (SAR) are broadspectrum plant defense responses that can be induced biologically by challenging a plant with a weaker strain of a specific pathogen or exposing a plant to natural and/or synthetic chemical compounds (Elliston et al. 1977). SAR has been studied by plant biologists for the past 100 years as a means to increase resistance to fungal, bacterial, and viral pathogens in crop plants such as potato, wheat, and rice (Agrios 1997).
The objectives of this review are to discuss the concepts of SAR to raise awareness of the potential and past uses of this technique to control plant diseases and, where applicable, highlight for urban foresters how SAR could be incorporated into a pathogen management system within towns and cities. It is important, however, to emphasize that SAR as a system to control tree diseases has received little scientific investigation (Karokene et al. 1999). Consequently, examples used throughout this paper rely heavily on results produced from herbaceous crop research.
HISTORICAL BACKGROUND
The natural phenomenon of resistance development in response to pathogen infection was initially recognized by Ray (1901) and Beauvene (1901), working with Botrytis cinerea (gray mold). Beauvene (1899) had previously discovered that the virulence of a strain of B. cinerea could be varied by pre-exposing the pathogen to heat or cold. He then induced SAR in Begonia, by planting in soil previously inoculated with the heat- or cold-treated strains or by injecting inoculum directly into the plant. Regardless of the inoculation technique used, plant resistance developed to subsequent infections with highly virulent strains of the same fungus. The first controlled laboratory study of SAR was performed by Ross (1961), who demonstrated that inoculation of a single leaf of tobacco with tobacco mosaic virus (TMV) reduced the severity of subsequent infections on other leaves. He coined the term “SAR” for the resistance that developed in the untreated portions of TMV-inoculated plants. Extending work on SAR to fungi, Cruickshank and Mandryk (1960) showed that high levels of resistance against Peronospora tabacina (blue mold) could be achieved within 3 weeks following inoculation in field-grown tobacco when a spore suspension of the same fungus was injected into stem tissue (Cohen and Kuc 1981).
Other recent, well-characterized examples of SAR have been recorded in both dicotyledonous and monocotyledonous plants (Hunt and Ryals 1996; Schneider et al. 1997; Mauch-Mani and Metraux 1998).
PERSISTENCE OF SAR
Many diseases and decay processes in woody tissue develop over many years; however, the effect of inducing an SAR response upon retarding existing disease infection within a tree is unknown (Christiansen et al. 1999). Once induced, the SAR response in spruce (Picea abies) has been observed to persist for at least 1 year and possibly longer following inoculation, leading to the interesting hypothesis that a sublethal attack one year may strengthen rather than weaken a tree’s defense (Christiansen, personal communication). Once induced, SAR can lead to long-lasting, broad-spectrum disease control. For example, inoculation of cucumber with Pseudomonas lachrymans or Colletotrichum lagenarium provided systemic resistance within a few days against 13 separate diseases, including those caused by fungi, bacteria, and viruses. A single SAR-inducing infection protected cucumber plants for 44 weeks, while a booster inoculation 2 to 3 weeks after the primary infection led to season-long, broad-spectrum resistance (Kuc and Richmond 1977). Whether such broad-spectrum resistance occurs in trees has never been investigated. Further research in this area is now warranted.
MECHANISMS OF SAR
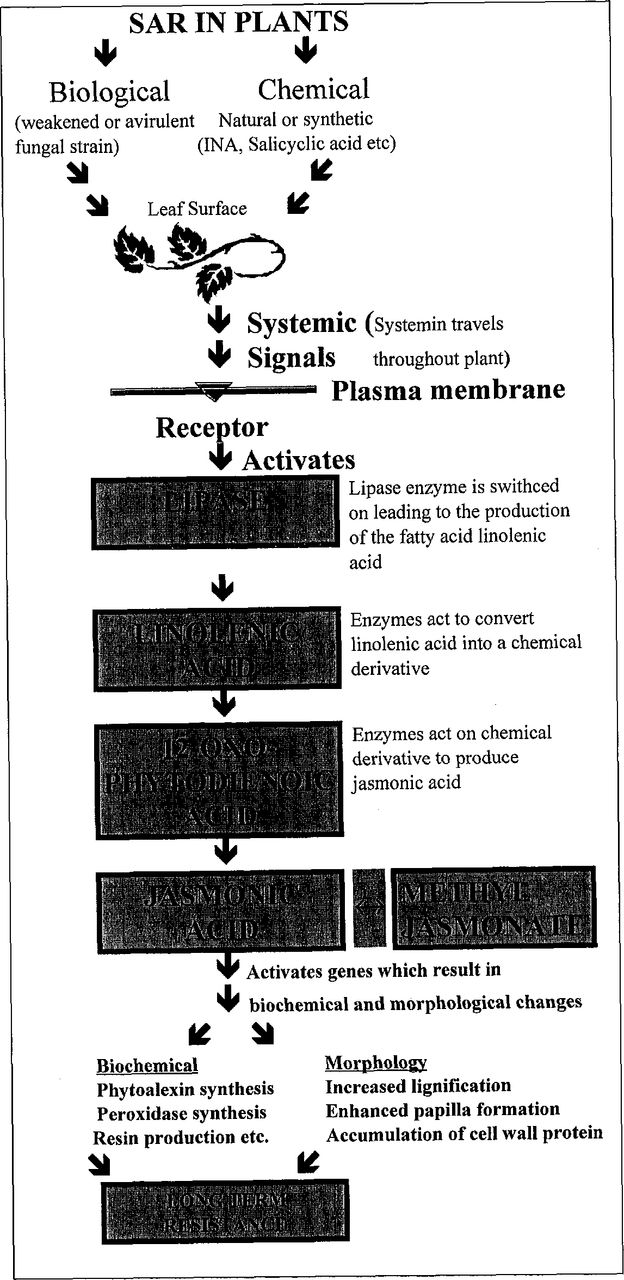
Van Loon and Van Kammen (1970) andGianinazzi et al. (1970) showed that viral infection of tobacco induced the accumulation of a distinct set of proteins, called pathogenesis-related proteins (PR proteins). Ward et al. (1991) demonstrated that at least nine gene families were induced in uninfected leaves of inoculated plants; these gene families are now known as SAR genes. Several of these SAR gene products have direct antimicrobial activity or are closely related to classes of antimicrobial proteins. These include β-1,3-glucanases, chitinases (Busam et al. 1997), cysteine-rich proteins related to thaumatin, beta-(l,3)-glucanase, and the PR-1 proteins (Anfoka and Buchenauer 1997). Further corroboration for the involvement of SAR genes in resistance comes from a range of transgenic plant experiments using seedling material (Alexander et al. 1993). The set of SAR genes that are induced differs among plant species. In cucumber, a class-III chitinase is the most highly induced SAR gene, while in tobacco and Arabidopsis, PR-1 and NPR1 are the predominant genes expressed (Cao et al. 1997, 1998). Such differences between species may reflect different evolutionary or breeding constraints that have selected for the most effective SAR response to the specific pathogen to which a species is most subject to attack.
Morphological and biochemical changes in SAR-protected plants that then become infected include a significantly faster lignification response, which corresponds with an increase in peroxidase activity (Ajilan and Potter 1992). Other changes include increased glucose and fructose concentrations in systemic tissue (Chandra and Bhatt 1998), an accumulation of fungi-toxic β-ionone derivatives (Wyatt and Kuc 1992), induction of lipoxygenase (Staub et al. 1992), antimicrobial fatty acid derivatives (Namai et al. 1993), phenylalanine ammonialysase, phytoalexins (Elliston et al. 1977), and hydroxyproline-rich glycoprotein (Raggi 1998). Within conifers, inducible defense systems include secondary resin production, synthesis of new phenolics, traumatic resin duct formation, and initiation of a wound periderm (Franceschi et al. 2000).
A schematic diagram of the SAR response, based on available literature, is presented in Figure 1. It is important to emphasize, however, that the exact nature of the long-distant systemic signal (systemin) has not yet been fully elucidated, and aspects of systemic signaling in SAR remain unknown (see Ward et al. 1991; Kessmann et al. 1994; Chen et al. 1994 for further information).

SAR response in plants.
SAR INDUCED BY BIOLOGICAL AGENTS
Although a range of organisms is commercially available for use as biocontrol agents, nearly all are based on a direct antibiotic principle. For an organism to be classified as an SAR activator, certain criteria need to be fulfilled (see section titled SAR Induced by Chemical Means). Consequently, whether the organisms discussed in this section can be classified as SAR activators is unknown and requires further research to confirm resistance-inducing activity. In all of these cases, therefore, mechanisms other than SAR may be involved in increasing resistance.
Kessmann et al. (1994) reported that an extract of Rheynoutria sachalinensis gave good control of powdery mildews and other crop pathogens. However, the active principle(s) are not known. Extracts from Bacillus subtilis have been reported to induce resistance in barley, especially against powdery mildew (Steiner et al. 1988), while an infiltration of Penicillum janczewskii or its culture filtrate into melon and cotton increased peroxidase activity (a response associated with SAR) and resulted in increased protection against Rhizoctonia solani and elimination of the incidence of dampening-off symptoms (Madi and Katan 1998).
Another interesting class of agents is plant growth-promoting rhizobacteria, which are able to protect plants against foliar pathogens when used as a seed soak treatment (Maurhoffer et al. 1994). This strategy offers an exciting potential since disease control and increased tree health can be combined in a single seed treatment with naturally occurring microorganisms. A report examined the induction of SAR by a Pseudomonas fluorescens biocontrol strain (Maurhoffer et al. 1994). They demonstrated that growing plants in soil inoculated with this bacterium induced symptoms of SAR in leaf tissue. This response was absent in plants grown in uninoculated soil, or in soil inoculated with a different non-SAR-inducing strain. Similarly, Wei et al. (1991) reported on strains of Pseudomonas able to protect cucumber against a range of diseases. Mercer and Kirk (1984a,b) reported a series of laboratory tests designed to select microorganisms that would be effective in reducing infection by decay fungi and their subsequent performance under longterm field conditions. From preliminary tests of 22 bacterial isolates, three isolates of Bacillus spp. were selected with antibiotic properties. Whether Pseudomonas fluorescens was one of the 22 bacterial isolates used was not reported; however, the use of Pseudomonas fluorescens as a dual SAR inducer and biocontrol agent may warrant further investigation. Inducing SAR by chemical means and simultaneously inoculating with Bacillus UW85, a non-SAR-inducing microbial biocontrol strain, to investigate how expression of SAR affected the efficacy of this biocontrol agent was undertaken by Chen et al. (1996). Interestingly, SAR reduced the mortality of tobacco seedlings inoculated with zoospores of fungal diseases but had no effect on the growth of UW85. Combination of SAR and treatment with UW85 resulted in additive suppression of disease development.
Verticilum dahliae, to be marketed as DutchTrig® (Arcadis, Inc., the Netherlands), is under current trial for the biocontrol of Dutch elm disease. Although promising, the practical use of microorganisms to induce SAR at present is most likely restricted to selected trees grown on a small scale under controlled or glasshouse conditions and has useful implications for the tree- and shrubgrowing industry. Interestingly, no commercial biological product with an SAR mode of action has been introduced into the market, although DutchTrig® offers a possibility for the future.
SAR INDUCED BY CHEMICALS
Chemicals that are able to induce SAR would offer a number of advantages over current conventional techniques for disease control in trees. Even if only a portion of the canopy is sprayed, the SAR response would be transduced throughout the canopy to provide long-term resistance. Similarly, unlike the classical biological control concept based on a predator—prey situation, SAR retains its efficacy irrespective of adverse environmental conditions such as low temperatures.
According to Kessmann et al. (1994), three criteria need to be fulfilled before a chemical agent can be classified as an “activator” of the SAR response: 1) the treated plants are resistant to the same number and type of diseases as those plants in which SAR has been biologically induced, 2) the chemical used has no direct antimicrobial activity or can be converted by the tree into antimicrobial metabolites, and 3) the same pre-infectional biochemical processes are induced as recorded in plant tissues after biological induction of SAR.
Salicylic Acid (SA)
Although salicylic acid (aspirin) is recognized as an inducer of PR protein accumulation and SAR resistance when sprayed onto plants, and it fulfills all of the criteria of an activator, there are a number of problems with using SA as an SAR-inducing chemical. Reports of SA-mediated resistance are restricted to effects in the treated tissue, indicating that SA does not translocate efficiently throughout the plants when applied exogenously (Enyedi and Raskin 1993). Similarly, SA which is deposited upon a leaf surface, is rapidly broken down—resulting in only a short-term response. Finally, only a narrow safety margin separates the rates at which the compound is effective and the rate at which it becomes strongly phytotoxic. Because of these limitations, SA has not been considered a practical solution to disease control.
INA
A far more promising chemical is 2,6-dichloroisonicotinic acid (CGA 41396) and its methylester (CGA 41397), both referred to as INA. These compounds were discovered as able to induce systemic resistance in plants (Jensen et al. 1998), which provided good protection against fungal and bacterial pathogens of crops under glasshouse and field conditions (Table 1). Importantly, whether applied as a foliar spray or root drench, the spectrum of SAR activity is identical to the resistance observed after local pre-infection with SAR-inducing biological agents.
Activity of GCA 41396 in field trials (after Kessmann et al. 1994).
Plant responses to INA and INA analogs include induction of β-1,3-glucanases, chitinases, 6-phosphogluconate-dehydrogenase, various phenylpropanoid-derived metabolites, accelerated lipid metabolism, and peroxidase(s) synthesis (Seguchi et al. 1992; Staub et al. 1992), resulting in high-level resistance against plant diseases within 2 days of application. Histological studies showed that plants treated with INA respond to infection by a single-cell necrosis at the site of attempted penetration. The few hyphae that managed to penetrate become surrounded by a cluster of necrotic cells and fungal growth ceases (Madamanchi and Kuc 1991).
Other Chemicals
A number of reports have claimed resistance-inducing activity for a range of chemicals or plant extracts. Although these compounds have failed to fulfill the criteria established for SAR activators, they still could be considered as part of an integrated disease management system.
Plants pre-treated with fosetyl-Al, metalaxyl (Ward 1984), and triazoles (Hauthal 1993) accumulate phytoalexins faster than nontreated plants. The use of triazoles to promote SAR may be of interest for urban foresters. Carbendazim is classified as a benzimidazole and forms part of the gel formulation Bayleton BM recommended for tree work under British Standard guidelines (British Standards Institute 1989). A range of fungitoxicants (furmecyclox, fenpropimorph, carbendazim, and triadimenol) applied as pruning wound treatments on apple trees following inoculation with Coriolus versicolor were investigated by Gendle and Clifford (1983). No SAR-inducing properties of these chemicals, however, were investigated, and in all cases mode of application was via small surface-area pruning wounds. Although speculative, if triazoles also induce SAR-type responses, application as a foliar spray to a larger surface area in conjunction with application to pruning wounds may prove more effective in disease control. Further research is warranted to confirm this hypothesis.
Benzo-(l,2,3)-thiadiazole-7-carbothioic acid S-methyl ester (BTH) and its derivatives induce SAR in a number of plants (Gorlach et al. 1996), while probenazole (Oryzemate®) is a systemic compound used successfully to protect rice against Pyricularia oryzae (Kato et al. 1984). Because of their tolerance and efficacy in crops, both compounds have been commercialized, but, surprisingly their use in urban forestry remains untested.
The use of ethylene-producing substances, such as Ethrel (ethephon), as a foliar spray to cause a timed and coordinated shedding of leaves to reduce street sweeping and cleaning costs has been suggested by Moore (1998). Similarly, such a substance could be used to manipulate a timed ripening and shedding of fruits that are undesirable or pose a potential hazard within an urban context. The ethylene-releasing agent ethephon has been shown to induce SAR gene expression (Kessmann et al. 1994). Any compound that can be used to manipulate plant growth and simultaneously promote an SAR response would obviously be of great benefit within an urban forestry situation.
Applications of low concentrations of jasmonic acid (JA) to plants induce proteinase inhibitors, proline-rich cell wall protein, and a range of enzymes involved in plant defense reactions (Sticher et al. 1997). Additionally, significant in vitro activity of JA has been reported (Schweizer et al. 1993). Use of JA within an urban forestry program may, however, have limitations since JA is also recognized as a growth regulator affecting a range of physiological processes including root growth, senescence of leaves, and stomatal opening (Moore 1998).
Other chemical or plant extract agents with proposed SAR-inducing activity include various inorganic salts, compost and compost water extracts, low molecular weight proteins (elicitins) silicon, oxalate, phosphate, 2-thiouracil, polyacrylic acid, nucleic acids, unsaturated fatty acids and N-Trimethyl-L-lysine (Kessmann et al. 1994).
INTERPLANTING WITH FLOWERING WOODY PLANTS
As mentioned above, spraying plants with JA or methyl jasmonate can induce an SAR response but presents a number of undesirable side effects. A novel route to immunization against tree pathogens, however, may be to continually expose plants to JA vapor. Indeed, horticulturists and amateur gardeners may already be using JA vapor to control plant pathogens without realizing it. Until recently, the concept of intercropping flowering plants within a glasshouse or small-scale field crop was believed to reduce pathogen and pest incidence as the flowering plant attracted a range of beneficial insects for pollination. It is now known that various species of Artemisia, for example, have high levels of JA in their leaves that slowly diffuse from leaf tissue into the atmosphere. Consequently, interspersing Artemisia within a crop of tomato resulted in induction of SAR and a significant reduction in disease incidence (Day 1993).
Planting an understory of woody plants (Berberis, Pyracantha, Mahonia, etc.) around a tree base is a recognized technique for suppressing weeds and simultaneously reducing the incidence of vandalism. The use of perennial flowering shrubs and/or short-lived flowering annuals as an understory may also provide a means of promoting an SAR response. Similarly, interplanting nonflowering trees with ornamental flowering species such as Prunus or Crataegus may induce an SAR response. With the exception of Jasminum spp., knowledge of flowering woody plants producing SAR-inducing chemicals within leaf, shoot, and flowering tissue is very limited but warrants further research. The results of a “desktop” literature search of plants possessing and on occasion diffusing JA from leaves, shoots, and flowers are shown in Table 2. Only plants suitable for use as an understory within existing tree plantings or those used by the landscape industry are presented.
Plants with potential for inducing SAR.
Because JA is a naturally occurring growth regulator, this compound will undoubtedly be found in a further range of plants not listed in Table 2. Further research is required to determine other plants that release SAR-inducing chemicals, concentrations released by these plants, and influences of life cycle (flowering, seed set, etc.) upon rates of release, as well as concentrations required to induce the SAR response in neighboring plants. Until further research provides answers to these questions, this technique, if feasible, is most likely restricted to trees grown on a small scale under controlled nursery or glasshouse conditions.
SAR IN WOODY PLANTS
Very little literature exists on the use of SAR in controlling diseases of woody plants. Of that available, however, the bulk has been undertaken by Christiansen and colleagues at the Norwegian Forest Institute, Ås, Norway, using Picea abies as models against the fungal disease Ceratocystis polonica and the Eurasian spruce bark beetle (Ips typographies). In summary, their results have shown that at least two different mechanisms are involved in the induction of enhanced resistance of P. abies. These are that polyphenolic parenchyma cells in the bark respond actively to wounding, and particularly to fungal infection, with wounds and beetle attacks in the outer phloem effectively enveloped and rendered harmless by these cells. Traumatic resin ducts are formed in the xylem of trees surviving I. typographies attacks or mass inoculation with C. polonica. These resinous materials produced by epithelial cells lining these ducts may contain newly formed phenolic material fungastatic in nature. Together the traumatic resin ducts and surrounding polyphenol-accumulating cells may provide a potent protective structure in which chemical substances accumulate and are ready to retard the longitudinal and inward spread of pathogenic organisms attempting invasion after wounding (Nagy et al. 2000). Induction of change in polyphenolic parenchyma cells were visible 6 days after induction, while traumatic duct formation required 2 weeks (Franceschi et al. 2000). Christiansen and Krokene (1999) concluded that “vaccination” with fungal inoculations can enhance the resistance of spruce trees to later beetle attack, although such “vaccination” of spruce trees is unlikely to have any practical application in forestry but may have potential for protecting valuable ornamental trees threatened by high bark beetle populations. Other work includes that of Okey and Sreenivan (1996), who recorded that application of SA as a seed treatment, spray, and soil drench, reduced the number of lesions, the size of the lesion area, and the percentage of necrotic leaf area of two clones of cacao infected by Phytophthora palmivora. Application of the SAR-inducing chemical INA as a foliar spray reduced fire blight (Erwinia carotovora) symptoms by 45% compared to noninoculated controls (Kessmann et al. 1994).
CONCLUSIONS
Historically, emphasis in plant pathology has been placed on the discovery of new resistance genes by breeding and molecular techniques rather than on using the resistance potential already present in plants. Recent demonstrations of the effectiveness of SAR in laboratory and field situations presents interesting opportunities for the control of plant diseases by the urban forester. Experiments with crop plants have shown that SAR can lead to long-lasting, broadspectrum disease control and can be used preventively to bolster general plant health. Ample evidence suggests that SAR is based on multiple natural defense mechanisms, and this makes it less likely that a pathogen can readily develop resistance to this control measure. The availability of this long-lasting, broad-spectrum and potentially stable solution to disease control may have an positive impact on urban forestry management.
- © 2001, International Society of Arboriculture. All rights reserved.
LITERATURE CITED
In this issue









{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.