
Abstract
We monitored crown air temperature, volumetric soil moisture, leaf water potential, rates of gas exchange, and insect pests of green ash (Fraxinus pennsylvanica ‘Marshall’s Seedless’), Austrian pine (Pinus nigra Arnold), and northern red oak (Quercus rubra L.) trees during a relatively hot summer in Lincoln, Nebraska, U.S. Air temperatures and vapor pressure deficits (VPD) were much higher in trees growing downtown than in nearby trees growing on the University of Nebraska campus. Increased VPDs and reduced soil moisture decreased pre-dawn water potential and gas exchange of ash and oak trees on the downtown site compared to trees on campus. Green ash trees downtown had more damage from lilac borers (Podosesia syringae (Harris) (Sesiidae)) than did trees on campus. Aphids (Aphidiae) and lace bugs (Tingidae Corythucha arcuata (Say)) appeared to be more numerous on oak trees growing downtown than on those growing on campus. Cultural treatments that improve soil moisture availability, such as irrigating, increasing planter size, and mulching, are especially critical on heat island sites and may reduce tree stress, pest damage, and mortality.
Urban tree planting programs are widely promoted as a means to reduce the effects of warming of urban environments caused by absorption, advection, and re-radiation of heat from streets and buildings, commonly referred to as the urban heat island effect. Temperatures in urban areas may be up to 8°C to 10°C warmer than in the surrounding countryside (Davidson 1988). Trees are effective at cooling microclimates through shade and transpiration. However, in urban environments trees face an array of man-made and natural stresses that may reduce their life spans compared to trees in rural areas or natural stands. Although estimates vary, life spans of trees in downtown areas are often less than years (Berrang et al. 1985; Gilbertson and Bradshaw 1985; Nowak and McBride 1991). Increasing our understanding of the relation of urban site factors to tree stress and health is critical for urban tree management, tree selection, and success of urban tree planting programs.
Some environmental stresses of trees in urban and suburban sites, such as acute exposure to de-icing salt (Fluckiger and Braun 1981) or environmental pollutants, are unique to those environments. However, other abiotic stresses, such as drought or extreme temperatures, occur in virtually all ecosystems in which trees are native but may be exacerbated by human development (Bassuk and Whitlow 1986). Trees may experience increased drought stress in urban environments due to reduced soil moisture availability (Berrang et al. 1985; Clark and Kjelgren 1989; Cody and Smith 1997) or atmospheric drought (Whitlow and Bassuk 1986, 1988; Whitlow et al. 1992). Numerous factors may lead to soil-moisture-related drought stress, including restricted soil volumes, reduced rainfall infiltration, and soil compaction. The extent to which soil moisture actually limits urban tree growth and development varies with site factors and, in some cases, soil moisture availability may be greater under pavement (Wagar and Franklin 1994). In a study of New York City street trees, Berrang et al. (1985) observed that excess soil water was a more frequent problem than soil moisture deficits.
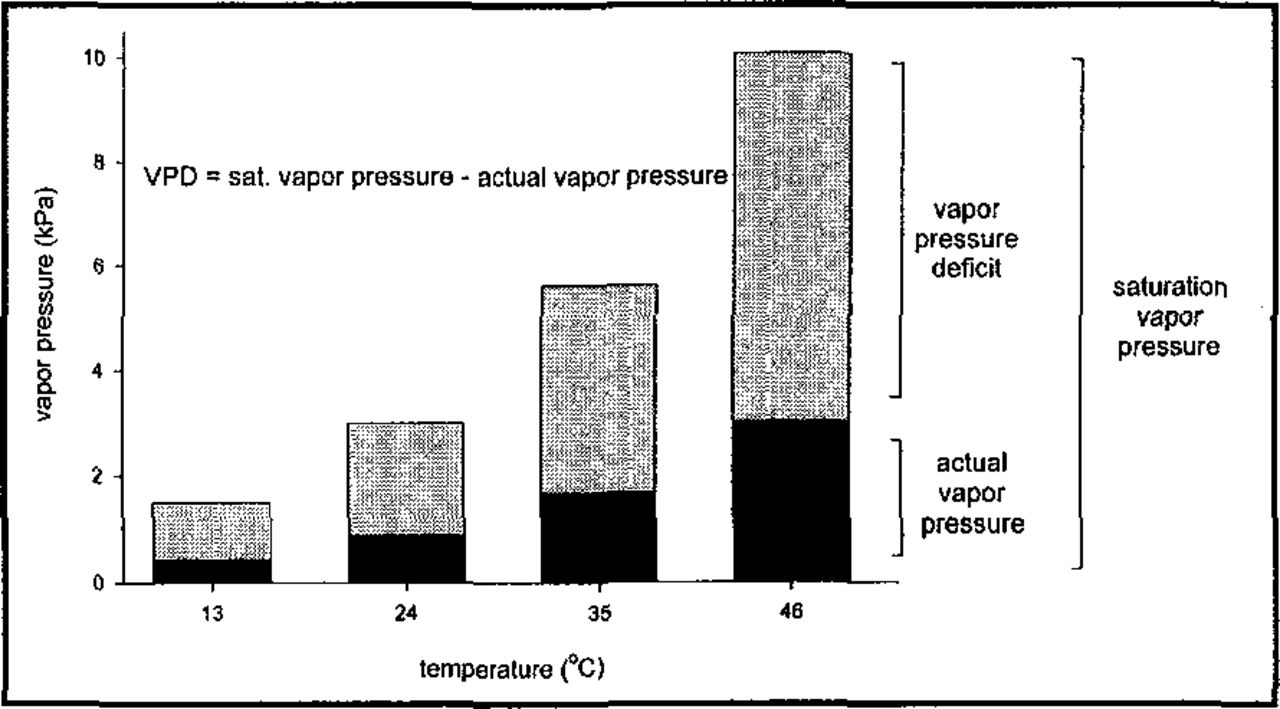
Atmospheric drought stress may occur when the rate of transpiration from tree crowns exceeds the capacity for roots to absorb water, resulting in decreased plant water potential (Kozlowski 1987). In considering the impact of urban heat island effects on tree water loss, it is important to understand that the “driving force” of transpiration is not just temperature but the vapor pressure deficit of the air surrounding the tree. Vapor pressure deficit (VPD) is calculated as the difference between the amount of water vapor in air compared to what the air could hold at saturation (saturation vapor pressure). Because saturation vapor pressure increases exponentially with temperature, VPD also increases exponentially with temperature at a given relative humidity To understand the potential impact of urban heat islands on tree water use, consider that at a relative humidity of 30%, each 11 °C increase in air temperature doubles the vapor pressure deficit (Figure 1). Therefore, relatively small increases in air temperature due to heat retention and re-radiation from buildings and asphalt can result in dramatic increases in evaporative demand and daily plant moisture stress.

Effect of temperature on vapor pressure deficit (VPD) at 30% relative humidity.
Increasing moisture stress can limit tree growth and reduce survival through direct and indirect effects on an array of physiological processes including photosynthesis (Cregg 1995), respiration, protein synthesis, and secondary carbohydrate metabolism (Kramer 1987). Moreover, reduced tree vigor caused by moisture stress may predispose trees to additional health problems, particularly insect infestation (Mattson and Haack 1987). The interaction of tree moisture status and insect damage is especially critical in the North American Great Plains because trees are often under moisture stress due to periodic droughts and because every tree species widely planted in the Plains is susceptible to at least one major insect pest (Dix et al. 1987). In order to quantify and better understand the effect of urban heat islands on tree microclimate, moisture stress, and susceptibility to insects, we studied paired groups of green ash (Fraxinus pennsylvanica ‘Marshall’s Seedless’), Austrian pine (Pinus nigra Arnold), and northern red oak (Quercus rubra L.) trees in downtown Lincoln, Nebraska, U.S., and in nearby stands on the East Campus of the University of Nebraska.
The objectives of the study were to
quantify the extent of the urban heat island effect between trees growing downtown and those growing on less-developed sites on campus;
compare plant water relations and gas exchange of three tree species on both sites;
compare insect pest abundance and insect damage between trees growing downtown and on campus.
MATERIALS AND METHODS
Site Description and Plant Materials
The study was conducted in the summer of 1995 in Lincoln, Nebraska. Lincoln is a city of approximately 200,000 people located in the central Great Plains of North America. The climate in Lincoln is continental; annual precipitation averages 685 mm (27 in.), the mean January minimum temperature is -13°C (8.6°F), and the mean July maximum temperature is 32°C (89.6°F). We studied paired plots of green ash, Austrian pine, and northern red oak trees. The plots were selected in order to have trees of roughly comparable size within a species at each location, although the downtown Austrian pines were considerably larger than the pines on the East Campus. At each site, we selected four trees of each species (Table 1). Within a species group, the trees were selected adjacent to one another to minimize time delays between gas exchange measurements and were representative of the overall planting. We measured initial tree height with a height pole and diameter at breast height (dbh) with a diameter tape. We estimated the age of each tree from increment cores collected at 0.6 m (2 ft) above ground. Trees at the East Campus were grown under open, park-like conditions. The ground under the trees was covered with turf, but the trees were not irrigated. The trees downtown were grown near the center of the main downtown business district in Lincoln, approximately 4 km (2.5 mi) from the East Campus study site. The downtown green ash trees were located on the south side of a three-story parking deck in sidewalk openings with a 1 m×1 m (3.3 × 3.3 ft) soil opening. The oak trees were located on the west side of the same parking deck as the green ash trees and were grown in raised planters with open bottoms approximately 2 m (6.6 ft) square and 0.6 m (2 ft) tall. The Austrian pine trees were grown in a parking lot adjacent to a large performing arts center 1 km (0.6 mi) from the green ash and oak plots. Austrian pines were mulched beyond the dripline with 5 to 7 cm (2 to 2.8 in.) of coarse pine bark. The other two species were not mulched.
Mean height and diameter and approximate age of study trees
Microclimate and Environmental Measurements
Air temperature and relative humidity were recorded hourly in the interior of the crown of one tree from each species at each location (six trees total) using portable sensors and dataloggers (Onset Computer Corp., Pocasset, MA). To shade and protect the dataloggers, we encased them in 7 cm (2.8 in.), white PVC end-caps with ventilation holes drilled in the bottoms. Official daily rainfall and temperature data were compiled from the National Weather Service Office (WSO) at the Lincoln Municipal Airport. We measured volumetric soil moisture at 15 cm (6 in.) and 30 cm (12 in.) near the base of each study tree using a TDR probe (Trase system, Soil Moisture, Inc., Santa Barbara, CA).
Leaf Water Potential and Gas Exchange
We measured leaf water potential (Ψ1), leaf conductance to water vapor (gwv), transpiration (E), and net photosynthesis (PS) on five dates in the summer of 1995. Leaf water potential was measured on single leaves or fascicles with a pressure chamber at predawn, late morning (0900-1100 h) and early afternoon (1300-1500 h) on June 20, July 6, July 13, July 27, and August 27. We measured gas exchange (E, gwv, and PS) at the same time as Ψ1 except for pre-dawn. Gas exchange was measured on detached shoots from three branches on each tree using an ADC LCA-4 portable gas exchange system (Analytical Development Co., Hoddeson, England). Preliminary trials indicated that gas exchange was stable for at least 10 minutes following detachment. We collected shoots from the mid- to upper canopy on each tree. Shoots from downtown green ash trees were collected from the upper level of the parking deck; all other shoots were collected using an extendable pole pruner. Using two or three-person work crews, we completed each measurement within 4 to 5 minutes of detachment. Although all gas exchange data were collected on clear to mostly sunny days, we used a portable light source to ensure that photosynthesis was measured under saturating light intensities (PPFD > 800 μmol m−2 s−1). To minimize rapid fluctuations in CO2 levels associated with nearby automobile traffic, inlet air for the portable photosynthesis system was supplied by a compressed gas cylinder (365-ppm CO2). The inlet air was humidified to approximate ambient relative humidity by a portable dew point generator (LI-610, Li- Cor Inc., Lincoln, NE).
Insect Surveys
We surveyed each tree at both locations for insect pests typically associated with the tree species in the central Great Plains (Dix et al. 1987). The oak trees were surveyed for aphids (Aphidiae) and lace bugs (Tingidae Corythucha arcuata (Say)), both of which are sap-sucking insects and may cause premature leaf drop. On seven dates during the summer, we collected four 15-cm-long (6-in.) shoots from each tree with a pole pruner and counted the number of aphids and lace bugs on four randomly selected leaves per shoot. We conducted a similar survey for the Austrian pine trees, sampling four randomly selected 15-cm-long shoots per tree and counting the total number of aphids on each shoot. The green ash trees were scored for lilac borers by examining the trunk of each tree to a height 3 m (10 ft) above ground. We counted the number of old (previous years’ damage) and active (indicated by fresh frass and fresh feeding scars) borer holes on each stem.
Analysis
Mean midday gas exchange rates and pre-dawn and midday Y1 were analyzed for site effects by repeated measures analysis of variance (Cody and Smith 1997). Means of insect number or damage were compared for each measurement date by unpaired t-test.
RESULTS
1995 Weather
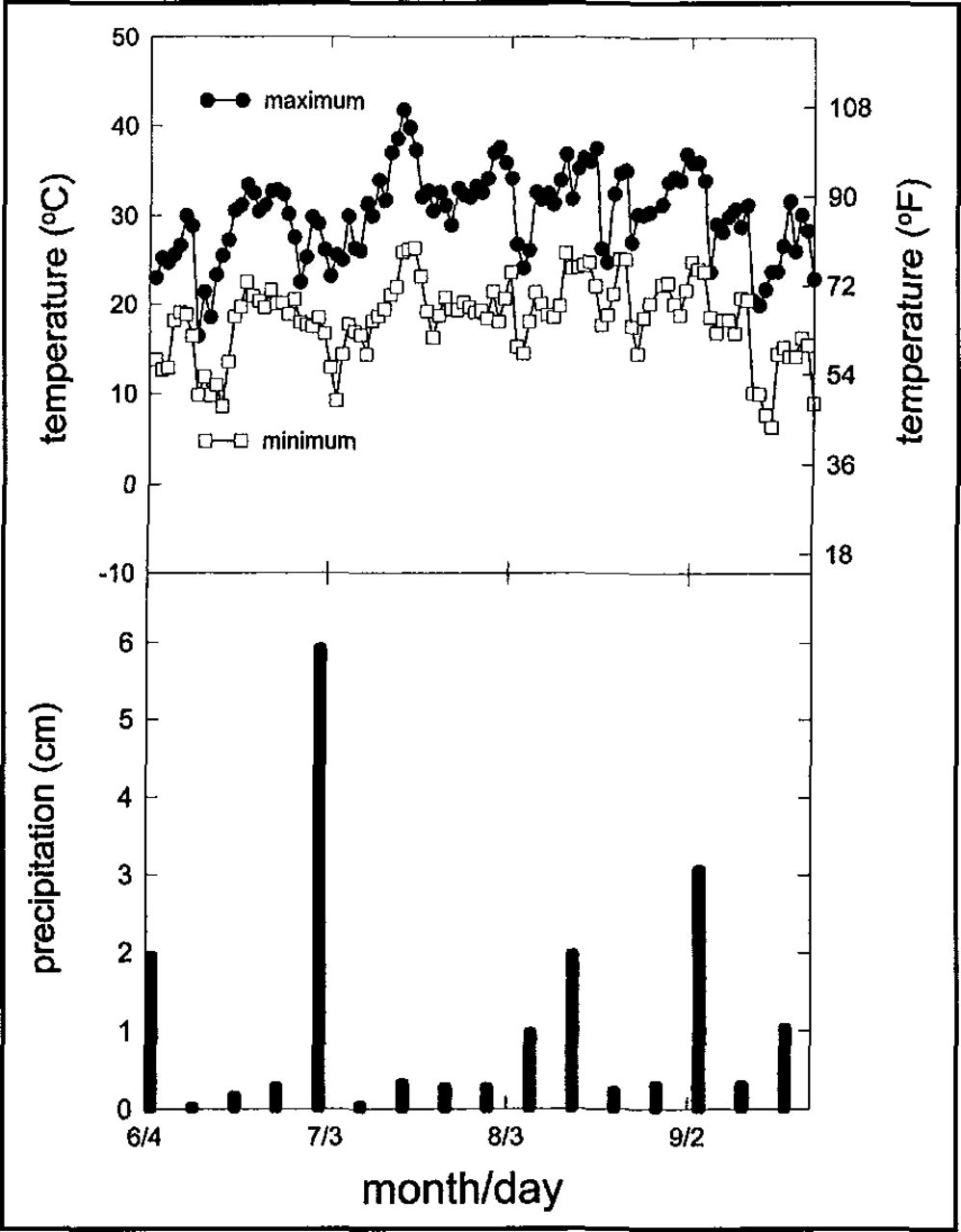
The weather during the summer of 1995 was one of the hottest in the region during the 1990s. The Lincoln Weather Service Office (WSO) recorded 10 days with maximum temperatures of 38°C (100°F) or higher, the most days over 38°C since 1988. Maximum temperatures at the WSO in Lincoln exceeded 38°C on five consecutive days in mid July (Figure 2). Following a heavy rain (3.7 cm [1.4 in.]) on the Fourth of July weekend, only 0.6 cm (0.2 in.) of rain fell in the next 30 days. Rainfall increased later in August and September.

Daily minimum and maximum temperature (top) and weekly precipitation (bottom) at the National Weather Service Office, Lincoln Metropolitan Airport, 1995.
Ambient air temperatures in the tree crowns were consistently higher downtown than on the East Campus. Through the July heat wave, mean daily temperatures were 3.3°C higher downtown than at the WSO, while temperatures on the East Campus averaged 1.4°C degrees warmer than at the WSO. The difference between microenvironments down-town and on the East Campus was most evident on July 12, the warmest day during the July heat wave (Figure 3). On July 12, the official high temperature at the Lincoln WSO reached 42°C (107.6°F). At the downtown study site the maximum temperature reached 52°C (125.6°F,) while the maximum temperature on the East Campus was 48°C (118.4°F). More significantly, the maximum VPD on the downtown study site (12.1 kPa) was 27% higher than at the East Campus location and almost double that observed at the WSO (6.3 kPa).
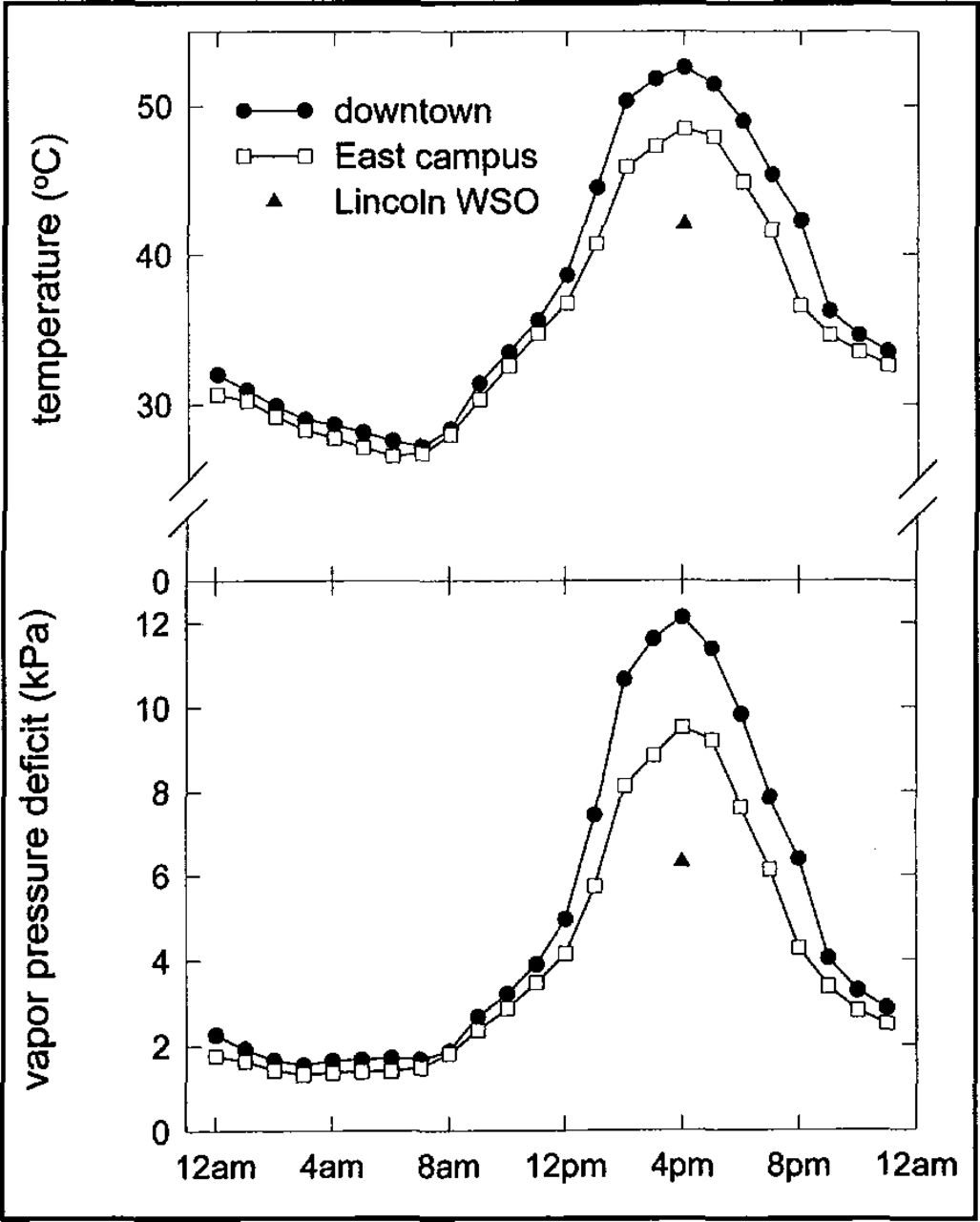
Mean hourly temperature and vapor pressure deficit at two locations in Lincoln, Nebraska, on July 12, 1995. Maximum temperature and VPD at the Lincoln Weather Service Office (WSO) indicated by ⯅.
Volumetric soil moisture differed significantly (P < 0.01) between sites and among dates (Table 2, Figure 4). At all locations, soil moisture peaked in early July and declined through the remainder of the study Except for initial soil moisture measurements in late June, soil moisture was consistently lower downtown than on the East Campus. Peak soil moistures on the East Campus exceeded 32% for all species. In contrast, soil moisture did not exceed 25% on the sites downtown. Among the species, the difference in soil moisture between the downtown and East Campus sites was greatest for the oaks. The lowest soil moisture during the study was 8.6% for the downtown oaks. Over the course of the study, soil moisture was 10.7% higher for the oaks on the East Campus than those downtown.
Analysis of variance for volumetric soil moisture (θ), pre-dawn leaf water potential (ψ1), and net photosynthesis (A) of green ash, red oak, and Austrian pine trees at two locations in Lincoln, Nebraska.

Mean volumetric soil moisture content of the upper 30 cm of soil near trees from three species at two locations in Lincoln, Nebraska, 1995. Note: Error bars indicate ± standard error of the mean.
Variation in soil moisture at the downtown site was reflected in tree water status during the study Predawn leaf water potential (Ψ1) declined with soil moisture following the rains in early July (Figure 5). For all three species, Ψ1 declined more rapidly on the downtown site than on the East Campus. In general, pre-dawn Ψ1, decreased rapidly once soil moisture approached 18% to 20%. Prior to the extended dry period in mid- to late July, there were no differences in pre-dawn Ψ1 between the downtown and East Campus sites, except for the Austrian pines that had slightly higher Ψ1 downtown than on the East Campus. Among the species, pre-dawn Ψ1, declined most rapidly for the hardwoods on the downtown site. Minimum midday Ψ1 (data not shown) did not vary consistently between sites. Minimum midday Ψ1 of the hardwoods ranged between -2.3 and -1.4 MPa, whereas minimum Ψ1 of the Austrian pines stayed within a relatively narrow range between -1.4 and -1.1.

Mean pre-dawn leaf water potential of trees from three species at two locations in Lincoln, Nebraska, 1995. Note: Error bars indicate ± standard error of the mean.
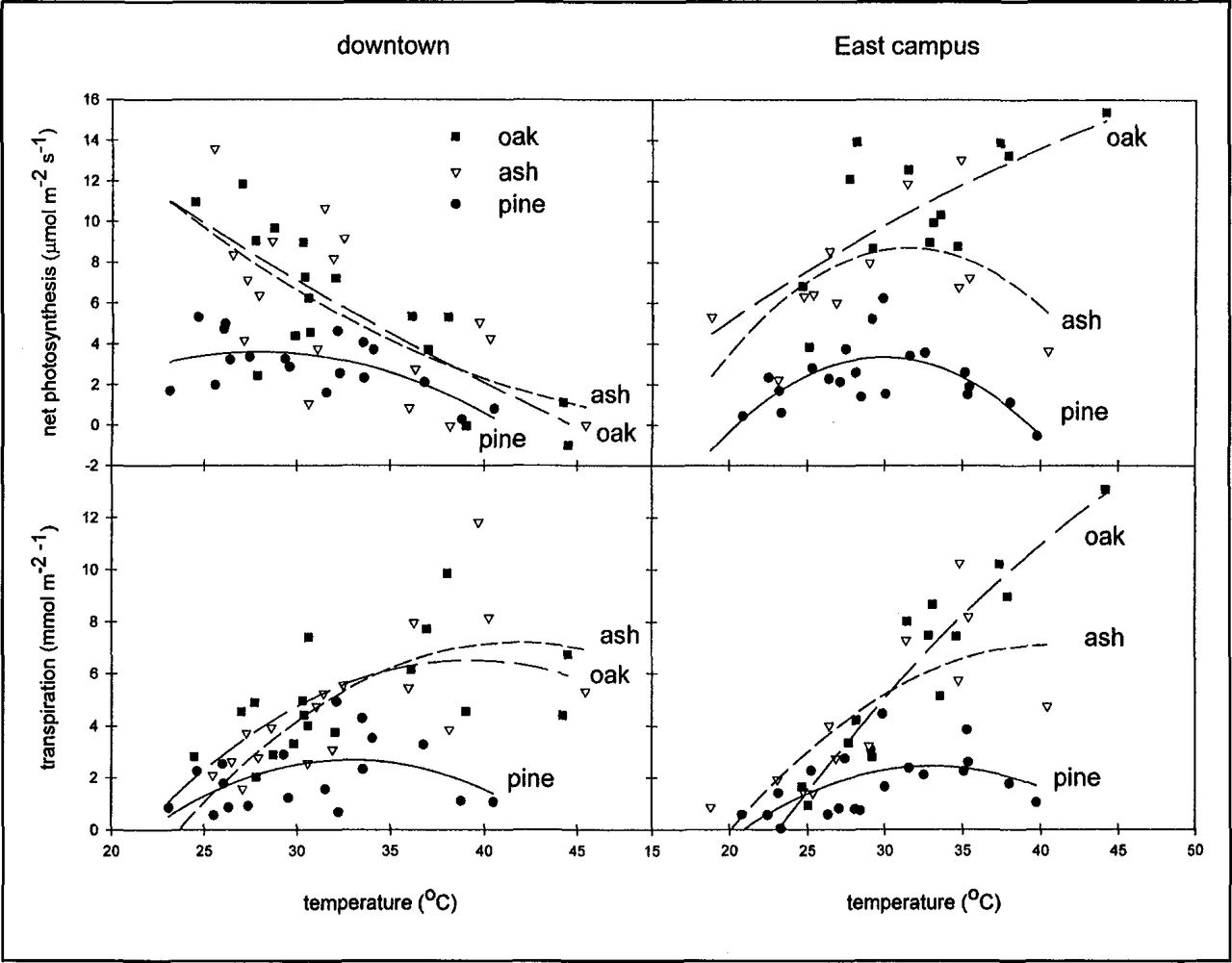
Variation in photosynthetic gas exchange was more complex than the seasonal trend in soil moisture and pre-dawn Ψ1 Overall, the oak trees had the highest rates of gas exchange and the pine trees had the lowest (Table 2, Figure 6). Differences in gas exchange between the two locations were greatest for the oak trees. In contrast, there was little difference in gas exchange in the Austrian pine grown at the two sites. The response of PS and E to increasing temperature (Figure 7) indicates that reduced soil moisture availability along with extreme temperatures combined to limit gas exchange on the downtown site. With increased soil moisture availability on the East Campus site (Figure 4), PS and E of the oak trees increased steadily with temperature. The water use and carbon gain response of pine trees to increasing temperature were similar at both sites whereas the response of green ash trees was intermediate between the oak and the pine trees
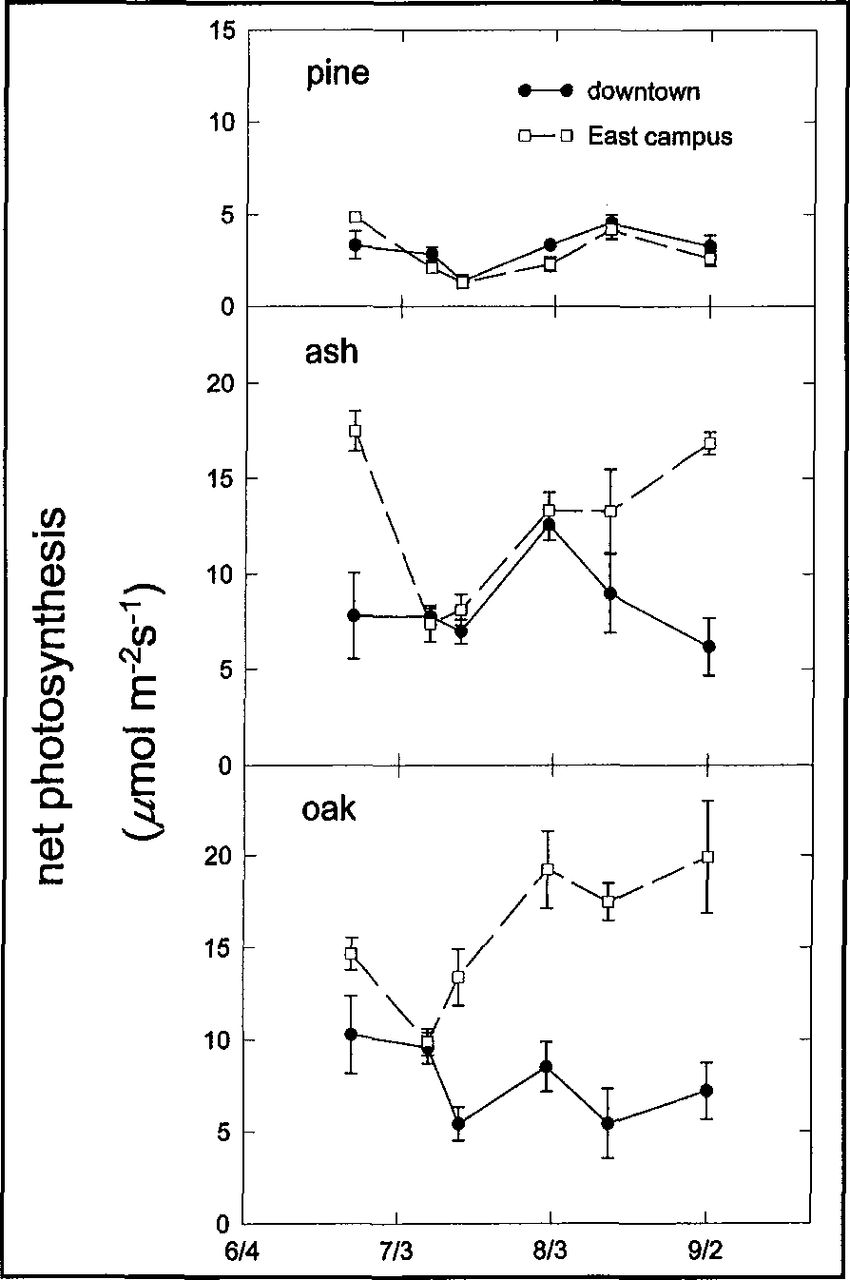
Mean midday (0900-1500 h) net photosynthesis of trees from three species at two locations in Lincoln, Nebraska, 1995. Note: Error bars indicate ± standard error of the mean.
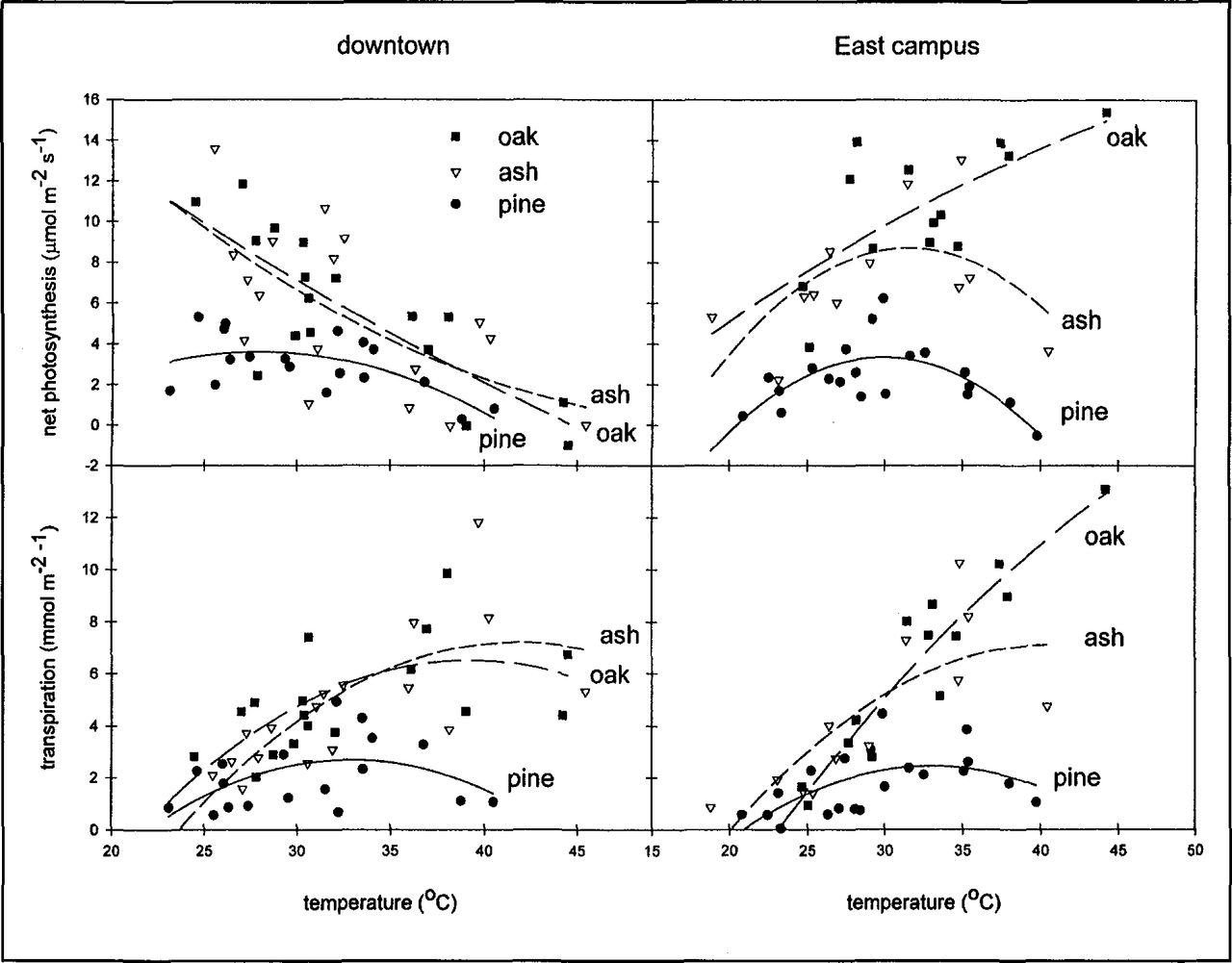
Response of net photosynthesis (top) and transpiration (bottom) to temperature of trees from three species at two locations in Lincoln, Nebraska, 1995.
Lilac borer activity was more common on green ash trees on the downtown site than on the East Campus (Table 3). Aphids and lace bugs appeared to be more numerous on the downtown oak trees than on the oaks on the East Campus, particularly late in the summer, but high variability prevented detection of statistical differences (Table 4). We did not observe any significant number of insect pests on the Austrian pines trees at either location during the study.
Lilac borer activity on green ash trees at two locations in Lincoln, Nebraska.
Survey of insect pests on northern red oak trees at two locations in Lincoln, Nebraska.
DISCUSSION
The results of this study demonstrate that it is difficult to overestimate the impact of urban heat islands on tree moisture stress and health, particularly for the hardwood species studied. Over the course of this study, increased tree moisture stress associated with downtown conditions reduced rates of net carbon gain of the oak and ash trees by 50% and 34%, respectively, compared to trees on the East Campus. The increase in tree moisture stress was related to a combination of above- and belowground factors. Soil moisture was generally lower downtown than on the East Campus. This reflects increased water use of the downtown trees associated with higher VPD and, perhaps, reduced rainfall infiltration and increased surface evaporation.
For the Austrian pines, soil moisture, pre-dawn Ψ1 and gas exchange were less affected by urban conditions than were the hardwood trees. The difference in the response of the Austrian pines compared to the hardwoods is likely related to several factors. First, the Austrian pines had much lower rates of gas exchange than the ash and the oaks when temperatures increased above 30°C (86°F). By maintaining relatively low rates of E, the pines were able to conserve soil moisture and thereby postpone the onset of drought stress. Secondly, unlike the hardwoods, the soil surface around the Austrian pines was mulched with 5 to 7 cm (2 to 2.8 in.) of coarse pine bark that conserved additional soil moisture by reducing evaporation from the soil surface.
The increase in lilac borers on the green ash trees was associated with the increase in tree moisture stress on the downtown site. Due to the confounding effect of geographic location, it is impossible to establish a causal relationship between increased moisture stress and insect infestation in this study. The relationship between moisture stress and insect attack is exceedingly complex and depends on a number of factors including type of insect, insect activity (e.g., feeding or ovipositing), portion of the plant affected (e.g., stem borers versus defoliators or sapsuckers), and the type of moisture stress (acute, short-term versus chronic, long-term). However, many previous studies suggest that, in general, moisture stress interferes with plant defense systems and increases the likelihood of insect attack (Mattson and Haack 1987). For example, infestation of mulberry trees by scale insects increased with increasing moisture stress and was higher on urban trees than forest trees (Hanks and Denno 1993). In contrast, Mopper and Whitham (1992) found that success of pine sawflies was often greater when trees were well-watered compared to drought-stressed trees. Thus, while the increase in aphids and lace bugs on the oaks may be due to increased moisture stress, it cannot be said with complete certainty.
The relationship between moisture stress and stem boring insects, such as the lilac borer, is clearer, however. With southern pine bark beetles and other pine bark beetles, moisture stress is the single most important factor in determining likelihood of infestation (Lorio and Hodges 1997). Thus, we speculate in the present study that the increase in lilac borer damage in the downtown ash trees is due to the increase in tree moisture stress at that location.
APPLICATION
There are several practical implications of the results of this study. First, to minimize moisture stress and reduce the likelihood of attack by insect pests, it is important to employ cultural techniques that increase or conserve soil moisture. Supplemental irrigation, where available, is the most direct means to increase available soil moisture and improve tree moisture status. However, as demonstrated by the response of the Austrian pines in this study, mulching can be very effective in conserving soil moisture and provides additional organic matter to the soil. Increasing planter size is another means of reducing tree moisture stress in urban settings (Cregg 1995). Lindsey and Bassuk (1991) have developed a useful model to estimate the planter size required for trees of various sizes.
Secondly, tree selection for urban environments must consider responses to increased temperatures and evaporative demand. Tolerance of soil compaction, road salts, and pollutants is frequently cited as a requisite trait for urban trees (Widrlechner 1994). However tolerance of atmospheric drought will also influence the degree to which trees experience moisture stress, and their ultimate longevity. Studies with forest trees indicate that tree species differ in their gas exchange response to VPD. For example, gas exchange of Pacific silver fir (Abies amabilis Dougl. ex Forbes) is very sensitive to increasing VPD (Martin et al. 1997), whereas gas exchange in loblolly pine (Pinus taeda L.) remains relatively constant across a wide range of VPD (Teskey et al. 1987). Additional data on the sensitivity of urban species to VPD could provide useful insights for urban tree selection.
Lastly, like many studies of urban effects on tree stress, this study is largely descriptive because the geographic separation between study trees results in the confounding of innumerable factors and precludes rigorous hypothesis testing. While this situation does not invalidate results, it points out the need for the development of well-designed urban tree or urban forest laboratories. Such sites should be dedicated to urban tree research and allow researchers to investigate effects of various urban stresses on trees planted in proximity and to develop a database of microclimate, growth, physiology, and health variables over time. These sites should be developed with an appropriate design to allow rigorous hypothesis testing and statistical analyses. As urban stress physiology research continues to come of age, developing urban stress laboratories will be the key next step to furthering the science of landscape tree selection and management.
Acknowledgements
This research was support by a grant from the International Society of Arboriculture Research Trust. We thank Rudy King, Bradley Rowe, and Carol Schumann for their review of an earlier version of this manuscript. We thank Darin Dauel and Scott Lee for their assistance with the field portion of the study.
Note: Use of trade names is for reader information and does not imply endorsement by the USDA of any product.
- © 2001, International Society of Arboriculture. All rights reserved.
In this issue









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.