
Abstract
Rowan (Sorbus aucuparia), a small and decorative tree with widespread distribution throughout Norway and Europe, is well suited for growth on street verges, parks, and urban woodlands. With a better knowledge of the genetic variation of its important traits, good genotypes could be selected and the appearance of the trees thus improved. The objective of this study was to describe the variation in growth, morphology, and phenology of rowan between and within seed sources and at different wind exposures. Seeds were collected from 10 to 15 trees within each of six populations in a small area of 1,300 km2 (500 mi2) along the southwestern coast of Norway. Three populations were wind-exposed and three were sheltered. From each of the 78 mother trees included, 30 half-siblings were planted in the field at the Særheim Research Centre (58.8°N, 5.5°E). Height growth, stem diameter, number of stems and branches, branch angle, and growth start and termination were recorded annually in five successive years. Large differences in most traits (estimated heritabilities varied from 0.14 to 1.0) were found among families within the stands. However, no important differences could be observed between progenies from wind-exposed trees and those from sheltered trees, and differences among stands were only minor. The results are discussed in relation to the climatic adaptation of rowan and how this plant material can be used in urban settings.
The visual and functional qualities of urban trees are largely dependent on the genetic quality of the plant material. While there is a need for both small and large trees, the larger trees often are too big for the available space, especially in gardens and paved areas.
Rowan (Sorbus aucuparia, L., 2n = 34, mountainash) is one of 12 Sorbus spp. growing naturally in Scandinavia, and eight of the species are endemic to the area (Salvesen 1993). Many Sorbus species are apomictic, but not rowan, in which nonapomictic seeds are set even after self-pollination (Sperens 1997). Rowan usually reaches a maximum height of between 8 and 15 m (25 and 50 ft) in Norway, but farther south in Europe the tree can reach a height of 30 m (100 ft). Its compound leaves have between 11 and 15 lancelot leaflets, and the white flowers, approximately 1 cm (0.4 in.) in diameter, have an inflorescence approximately 15 cm (6 in.) in diameter. Pollination is by insects and seed dispersal primarily by birds. The delicate leaf structure, the white and large inflorescence, the yellow to deep red fruits and often striking autumn colors are characteristics that make this moderately sized species valuable as an amenity tree in urban settings.
Rowan is widely distributed, from the mountains of Asia Minor to Siberia. It tolerates a wide range of climatic conditions, both continental and oceanic, and grows well in the harsh climates of high elevations (Hillebrand and Rosenberg 1996) and in sulfur-polluted environments (Nebe and Opfermann 1998). In Norway, it grows from the coast to elevations more than 1,500 m (4,900 ft) above sea level, where it is a shrub. Rowan thrives in many soil types, even in soil with poor nutrient status, but it grows best on rich farmland. Its excellent hardiness and large plasticity also make the tree suitable for use in urban woodlands, in sheltering belts, along streets, or in parks. Some of the trees form many stems, but others have only one. The single-stemmed type is preferred in paved areas, whereas in a sheltering belt, a tree with several stems might be more functional.
Improvement of the plant material for urban areas is necessary if amenity values and establishment are to be improved and maintenance costs decreased. The primary selection criterion for plants in Norway is climate adaptation, but other important criteria are growth potential, morphology, disease resistance, and decorative value. In urban woodlands, a plant’s ability to regenerate from seed is important.
Håbjørg (1978) has shown the significance of photoperiod as a signal of the termination of growth in the autumn and the preparation for winter hardiness in important tree species in Scandinavia. The rowan, however, has been found to be less dependent on the photoperiod for growth cessation, although the results are not conclusive. Selection programs can be designed depending on the importance of the species and available funding (Lagerstrüm and Eriksson 1996), but even in simple selection programs, knowledge of the genetic variation of important traits and distributions among and within populations is crucial for the design and success of the work.
Although some self-pollination occurs in rowan, the proportion of vital seeds is smaller after inbreeding than after outbreeding (Sperens 1997). Differences in crown density, fruit yield, fruit color, and chemical content of fruits have been found in rowan of different populations along a north to south gradient (Popov 1990). Phenological differences between populations of rowan have been interpreted as ecotype formation in rowan (Hillebrand and Rosenberg 1996). The genetic diversity in 17 European populations of rowan is high, both at the species level and within populations, whereas degree of differentiation is very low among populations (Raspe and Jacquemart 1998). Large within-stand variation, combined with little variation among populations, has been observed in beech (Konnert 1995; Larsen 1996), oak species (Kremer and Petit 1993; Bacilieri et al. 1994; Samuel et al. 1995; Herzog 1996), cherry (Frascaria et al. 1993; Mariette et al. 1997), and chestnut (Frascaria et al. 1993). However, in other studies, differences between populations have been found in beech (Löchelt and Franke 1995) and oak (Jensen 1993). There can be differences in the magnitude of genetic variation due to the pollination ecology of the species (Bruederle et al. 1998).
A selection program for rowan was started in Norway in 1993 with the objective of studying the genetic variation of progenies from different populations in a relatively small area along the southwestern coast. Also tested was whether trees growing on wind-exposed sites produced progenies different from the offspring of trees growing in sheltered sites. This paper presents data from growth, phenology, and morphology studied in a progeny trial at the Særheim Research Centre in Norway (59°N, 5.5°E).
MATERIALS AND METHODS
Seeds were collected from 10 to 15 trees within each of six stands in a small area of 1,300 km2 (500 mi2) along the southwestern coast of Norway. Three of the stands were wind-exposed and three were sheltered. A total of 78 mother trees were found, mainly along roads and stone walls in the cultural landscape, with fair to intensive agriculture. The seeds were washed out of the fruits after the harvest and sown in flats with a medium comprising 75% fertilized and limed peat (Floralux, Nittedal torvindustrier A/S) and 25% perlite. A thin layer of sand was applied as a topdressing to prevent loss of humidity from the stratification medium. Rowan has a long period of seed dormancy, so seeds were stratified in a cold store at 4°C (39.2°F) for five months. In March and April 1994, the flats with the stratified seeds were transferred to a greenhouse for germination. In May 1994, seedlings were potted (1 L [0.3 gal] root volume) for growing-on in the greenhouse until late June, when they were planted in the field. Three plants from each mother tree were planted in noncontiguous plots in ten randomized blocks (replications) in a well-drained Moraine soil. The distance between plants was 1.2 × 1.2 m (3.9 × 3.9 ft). Weed control was primarily done mechanically Data on growth, morphology, and phenology were collected in 1995, 1996, 1997, and 1998. The characteristics recorded were height, number of stems and branches, stem diameter, time and stage of leaf fall** on different dates (stages 1-9; 1 = 0%, 5 = 50%, and 9 = 100% of the leaves fallen), stages of budbreak* at a given date in 1996, 1997, and 1998 (stages 1-9; 1= no activity, 5 = leaves out of the bud scales but individual leaflets not visible, 9 = all leaflets are free and expanded), the general condition of each plant* (classes 1-9; 1 = unacceptable, 5 = average tree, 9 = excellent tree), diseases* (classes 1-9; 1 = no disease, 5 = some disease symptoms on 50% of the branches, 9 = the whole tree with disease symptoms), and winter damage* (classes 1-9; 1 = no damage, 5 = 50% of the branches with damage, 9 = all branches damaged). The characteristics marked with a single asterisk were recorded on a single date and those with a double asterisk on different dates.
The data were analyzed with the GLM procedure in SAS (SAS Institute, NC, USA) using a nested model. The effect of wind exposure was fixed, and populations within wind exposure and families within populations were treated as random components. Synthetic F-tests were used to test the differences between wind exposure and population. The residual error was used to test family variance.
The categorical data were transformed to normal scores based on midpoint values of cumulative frequency distributions of plants in different categories in each block (Gianola and Norton 1981; Falconer 1989; Ericsson 1994). The transformed data form a normal distribution with an expectation of 0 and a variance of 1 within each replicate.
Narrow sense heritabilities for individual progeny were calculated according to the approximation given by Squillace (1974) for open-pollinated families:
 where h2 = individual heritability in the narrow sense, σf2 = the variance among half-sib families, and σe2 = the environmental variance (residual variance). The variance components were estimated using the VARCOMP procedure (maximum likelihood method) in SAS. The standard errors of estimated heritabilities were calculated with the approximation given by Becker (1984):
where h2 = individual heritability in the narrow sense, σf2 = the variance among half-sib families, and σe2 = the environmental variance (residual variance). The variance components were estimated using the VARCOMP procedure (maximum likelihood method) in SAS. The standard errors of estimated heritabilities were calculated with the approximation given by Becker (1984):

RESULTS
Exposure and Populations
No differences were found in growth, morphology, or phenology between progenies of trees from different exposures. However, there were near-significant differences between plants of different exposure in the number of stems (P < 0.06) and the branch angle (P < 0.05), but the differences between the mean values were small (data not shown). There were also minor differences among the population means. However, statistically significant differences between populations were observed in the number of branches (P < 0.05), the growth form (shrub-like or tree-like, P < 0.05), branch angle (P < 0.001), and in time of leaf fall (P < 0.001) (data not shown). The differences among populations (in leaf fall) were observed only during the start phase of leaf fall, i.e., between 16 and 24 October. From 31 October to 7 November, no differences could be found (Figure 1a). The population means of height growth were almost significantly different (at P = 0.06–0.08 for 1995 through 1998).

Leaf loss on different dates for the populations (a) and some of the families (b), stages in terminal bud formation in the autumn of 1997 (c), and stages in budbreak (d) in spring 1998.
Families
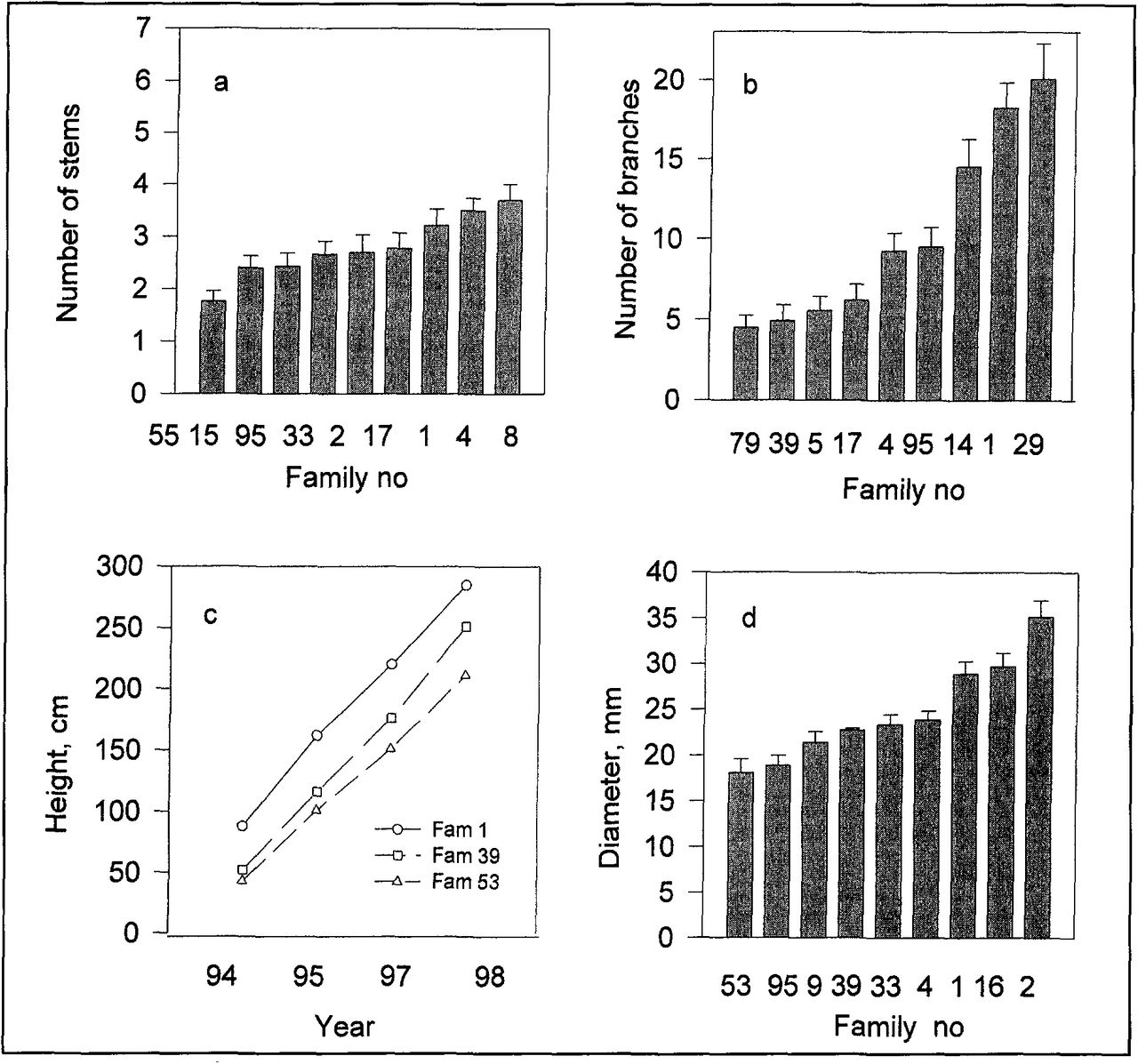
The differences among families were statistically significant (P < 0.001) for all recorded characters. The family means varied from 1.8 to 3.7 stems per tree (Figure 2a), a difference of 106%. The difference in mean number of branches was 347% (Figure 2b) between the families with fewest number of branches and those with the highest. After five growth seasons, trees of the smallest family were 211 cm (83 in.) and of the tallest 290 cm (114.3 in.), a difference of 37%. However, this difference was proportionately greater at the start of the field experiment, with a difference between highest and lowest family of 196% (49 cm [19.3 in.]) after the 1995 growing season (Figure 2c). The family with the smallest mean diameter had 94% less diameter than the family with the largest (Figure 2d) after the 1998 growing season.

Number of stems (a), number of branches (b), tree height (c), and diameter of the main stem (d) in some rowan families. The values are means ± standard error, n = 28-30.
Leaf fall began in the period from the end of September to early October. There was a statistically significant difference in time of shedding leaves, i.e., about a week between the earliest and the latest families (Figure 1a,b). The difference in stage of terminal bud formation, in August, was greater (Figure 1c), with some families having only 27% of individuals with a terminal bud and others with 100%. There was also a difference between families in time of budbreak, classified on a scale of 0 to 9. On 24 March 1998 (Figure 1d) some families were scored to have almost no development and some had reached the stage when the bud had started to show green in color. In 1997, the same evaluation of phenological development was made on a slightly later date, and the variation was from some development, but no visible green color, to stages where the individual leaflets were expanded (data not shown).
Heritability (h2) was fairly high for several of the phenotypic traits studied. For height, it was large, especially the first two years (h2 = 0.46 ± 0.09). However, it decreased to 0.36 and 0.28 in the third and fourth growth seasons. High values of heritability were also found for diameter (0.40 ± 0.08), number of branches (0.40 ± 0.08), growth start (0.65 ± 0.12, 1.0 ±0.17, 0.94 ±0.16 in 1996, 1997, and 1998 respectively), and time of leaf fall (0.28 ± 0.06, 0.42 ± 0.08, 0.48 ± 0.09 in 1996, 1997, and 1998, respectively).
Phenotypic Correlations Among Family Means
A positive correlation was found between diameter of the main stem and height of the trees (P = 0.001, R2 = 0.79). The families with the highest values for mean height tended to have the highest number of branches (P/R2 = 0.001/0.63). In the evaluation of the general condition of the trees (scored on a 0–9 scale), there was a correlation with height in the first three years, with a decrease in the strength of the correlation (P/R2 = 0.0001/0.56, 0.0001/0.50, and 0.003/0.34 for 1995, 1996, and 1997, respectively). There was also a correlation between the general condition of the families and the mean number of branches (P/R2 = 0.0001/0.42). There were significant correlations, although weak, between height and start of growth and time of leaf drop. Some other correlations were also statistically significant, but the P-level and the R2-values showed weak correlations, and the results are not presented. However, the lack of correlation between the formation of terminal buds and time of leaf fall is worthy of some attention.
DISCUSSION
Effects of Wind Exposure
The large family variation within each population indicates the presence of genetic variation. Thus, maladapted genotypes could be eliminated by strong winds, thereby creating differentiation among wind-exposed and sheltered populations in the traits selected against. It is commonly observed that exposed trees are formed morphologically by the predominant wind directions near the coast in Norway. The site at Særheim is exposed to wind, but mortality in the field trial has so far been low, with the mean differences between the two groups small and mostly nonsignificant for the traits measured. This indicates that the rowan expresses phenotypic plasticity by attaining different phenotypic growth forms from the same genotypes in response to differences in wind conditions.
Variation Among Populations
The collection of genotypes in the present study is not representative of sufficient gradients of latitudes or altitudes to determine the genetic variation among populations for adaptive traits related to fitness under different temperature conditions. So the lack of differences in growth and development among the populations may not be surprising. Even when statistically significant differences among the populations were observed in budbreak and time of leaf drop, they were small. Large within-population variation and smaller among-population variation are found in many tree species (Frascaria et al. 1993; Kremer and Petit 1993; Bacilieri et al. 1994; Konnert 1995; Samuel et al. 1995; Herzog 1996; Larsen 1996; Mariette et al. 1997; Raspe and Jacquemart 1998). Birds disperse the seeds of rowan, and so the gene flow among neighboring populations increases the within-population variation and counteracts the process of population differentiation.
Variation Among Families
The heritability estimates indicated a great deal of genetic variation in the sampled material. Open-pollinated families may contain a significant proportion of full-sibs within each family. The normal assumption that the variance component based on half-sib family means expresses 25% of the total additive genetic variation in a population (Falconer 1989) may therefore lead to an overestimation of the additive genetic component. Variance components based on full-sib family means are assumed to comprise 50% of the additive and 25% of the nonadditive variation in a population. A proportion value between these two situations is reasonable for use, and that is why an approximation of 33% was chosen (see Materials and Methods). However, this is based on experiences with wind-pollinated species. Rowan is an insect-pollinated species, and the proportion of full-sibs in an open-pollinated family may thus be even greater than in a family of a wind-pollinated species. One of the estimates of heritability in flushing in spring was actually greater than 1 (in 1997), and estimates were fairly high for growth and diameter. This could imply that the expression used is a slight overestimate of the genetic variation. Nevertheless, the genetic variation is large enough for a breeding program to be designed for rowan.
The relevance of reporting the growth and development of trees after only five years of field-testing may be questionable. However, in a compilation of results from the literature, juvenile-mature correlations were found to be good after only four years’ testing of trees in Pinaceae (Lambeth 1980). With the short juvenile phase of rowan, a positive result from early selection is probable and relevant in breeding. There may be several reasons why difference in height between the families decreased. From observations in 1998 and 1999, there seems to have been an increase in the proportion of trees that form flower buds. The height growth of rowan plants may therefore begin to diminish after the first six to eight growing seasons, because of the transfer from vegetative to generative growth, which is known to decrease the vegetative growth. The time of transfer from juvenile to adult stage might also be dependent on the climate and nutritional factors in the plant environment. In the wind-exposed test field, the highest trees will be most affected by the wind compared to those growing in shelter among the other trees.
Some families had a high proportion of plants with multiple stems and shrub-like growth. Some extremely shrubby individuals were also found, these being among the lowest, with a wide and open growth form and with branches almost hanging in some cases. These trees may be the result of self-pollination, with a decrease in vigor in the progeny.
The lack of correlation between formation of terminal bud and time of leaf drop may indicate that time of leaf drop is not a good predictor of growth termination and buildup of frost hardiness. The start of growth differed between families, but by the first week in May, most of the trees of the populations had reached the stage of expanded leaves. At the site of this study, the leaves rarely remain on the trees long enough for good autumn color to development. They are usually blown off in a storm before this. In the present study, the leaves were gone by mid to late October. The period with active photosynthesis is therefore about 150 days. Early budbreak and late growth termination correlated with height growth in birch (Wang and Tigerstedt 1993). This is in agreement with the present study, although the correlation in the present study was weak. For evaluation of hardiness in the autumn, a freeze test would probably have been more appropriate. Frost injury to the trees was actually minimal, if at all. The species is known to be very frost tolerant (Hillebrand and Rosenberg 1996).
Breeding for Urban Settings
The variation in the plant material of the present study clearly shows there is a potential for breeding for most of the characters recorded. This probably holds for many of the species of interest to urban planners and landscapers. Earlier breeding in trees focused primarily on species important for forestry Important selection criteria have been growth and wood and stem quality, which are not necessarily the most interesting traits for trees in the urban environment. Instead of maximum growth and size, appropriate growth, longevity, and resistance to pests and diseases may be characteristics of greater relevance. In rowan of the present study, in addition to the climate adaptation, the number of stems and the overall morphology may be characteristics of special interest in a breeding program for urban trees.
The importance of urban forests and trees is increasing, and the quality of green areas has to be given more attention. Breeding programs should be started to find healthy and high-quality progeny of important species for urban greenspaces, and the first step should be to identify the most important species and the selection criteria, which may differ depending on the site where the plants are to be used. The value of good individual plants may be higher in an urban context than in forestry, so clone propagation or selection of good families and the establishment of plantations for the production of propagation material may justify higher costs. With an international emphasis on selection in some important species, the possibilities of improving the quality of trees will probably result in a higher quality of greenspaces and less expenditure on their maintenance. Breeding in species for urban settings may thus lead to more and better greenspaces for the same money as spent today.
CONCLUSIONS
Seed harvest should not take place too far away (latitudes and altitude) from where the plants are to be planted. However, the individual mother tree seems be more important for the quality of the progeny than the population. A test of the progeny should be done before mother trees are used as seed sources.
- Copyright © 2000, International Society of Arboriculture. All rights reserved.
In this issue









{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.