
Abstract
Physiologic measurements of the internal water relations of sugar maple in a natural forest and along an urban street in Michigan were performed as a complement to a companion site characterization study. Investigations were carried out to determine how street trees are affected by the urban habitat, and to confirm through correlation analysis that site-related water stress adversely influences tree growth and vitality. Significant water relations differences exist between the street site sugar maples and those in natural habitats. Predawn water potential, osmotic potential, and stomatal conductance were significantly lower in the street trees, and were significantly correlated with the low soil moisture levels and high atmospheric demands of the street site. Findings of this study, in conjunction with the site characterization data, can be of value in selecting appropriate remedial treatments for stressed urban trees, as well as to reiterate the need for further screening of drought-tolerant cultivars for urban sites.
Erratum
In Urban vs. Natural Sugar Maple Growth: I. Stress Symptoms and Phenology in Relation to Site Characteristics, Journal of Arboriculture 22(3): 144-150, the charts in Figure 1 and Figure 2 were inadvertently transposed. The captions for those figures are correct.
This study was carried out in conjunction with a profile comparison study of tree growth, phenology, and site characteristics reported in Part I (3), which was conducted using urban street and natural forest sugar maples (Acer saccharum) in Michigan. Results of that comparison indicate that water stress may be the most important factor adversely influencing the growth and vitality of the campus street trees located at Michigan State University in East Lansing. Natural forest comparison data were obtained from the Minnis site, located 1.6 km from the campus street site.
This study was developed to compare sugar maple physiology in the natural forest and in an urban street site in order to confirm the impact of water stress, brought on by site conditions, on the growth processes of the campus street trees. Physiologic data quantify specific components of tree water relations, including water potentials, transpiration, and stomatal conductance. These are all controlled by environmental site factors such as soil water supply, relative humidity, and air temperature.
With the data from the companion site characterization study and from this water relations study, correlations can be made with the environmental site factors that most influence water stress in street trees on the campus site. Comparing these results with Minnis forest trees, as well as with other sugar maple studies in forest sites, enables the determination of differences in the adaptation of sugar maple to its environment when placed in an urban habitat.
Methods and Materials
The study sites are the same as those used for the companion site profile comparison study at Michigan State University (3). Seven trees were selected from each of 2 sites: an urban street, referred to as the campus site, and a natural forest, referred to as the Minnis forest. At the campus site, 7 sugar maples with diameter-at-breast-height of 12 to 20 cm were selected. All street trees were growing in a tree lawn of 9-m width. Seven dominant, naturally regenerated sugar maples were also selected in the Minnis forest. All study trees were free of visual decay symptoms, damage, and dieback.
Predawn Water Potential and Midday Water Potential
Water potential, measured in a leafy shoot, is a measure of tree water stress. Water potential was determined on 3 twigs from the mid crown of each tree, using the pressure chamber technique (13). Water potential measurements were taken on 6 dates during the summer of 1993. Predawn readings were taken between 4:00 and 6:00 A.M. Midday readings were taken between 12:00 noon and 2:00 P.M. from sunlit leaves.
Stomatal Conductance and Transpiration
Stomatal conductance expresses the regulatory control exerted by leaf stomata on transpiration rates. The greater the degree of closure, the less water vapor that flows from the leaf via transpiration, thus conserving plant moisture. Data were obtained using a Li-Cor LI-1600 Steady State Porometer on 3 sunlit leaves in the mid crown of each tree. Measurements were taken at midday between 12:00 noon and 2:00 P.M. on the same dates as, and in conjunction with, midday water potential readings.
Osmotic Potential
This variable is associated with the amount of water contained in the leaf cells. Low relative values (more negative) are indicative of water deficits occurring in the tree. Leaves were excised and gathered from the mid crown of each tree in conjunction with the 6 water relations measurement dates in July through September. Leaf sap was processed through a Wescor 5500 Vapor Pressure Osmometer to determine osmotic potential.
Data Analysis
Analysis of individual factors was performed using the nonparametric, two sample Mann-Whitney Rank Sum Test of the Systat statistical software package (SYSTAT, Inc., Evanston, Illinois). Results were judged significant and highly significant when probability levels were less than or equal to 5% (0.05) and 1% (0.01), respectively.
Correlation analysis was utilized to determine relationships between the tree water relations components and environmental factors on each site. The factors examined were the relationships and influence of the environmental variables on water potential and stomatal conductance. Data on the environmental factors, soil moisture at 15 and 30 cm, air temperature, and vapor pressure deficit were presented in Part I (3).
Single and multiple linear regressions were performed on the pooled data considering predawn water potential, midday water potential, and stomatal conductance, each separately as the dependent variable, and the environmental factors, considered separately and in combinations as the independent variables.
Results and Discussion
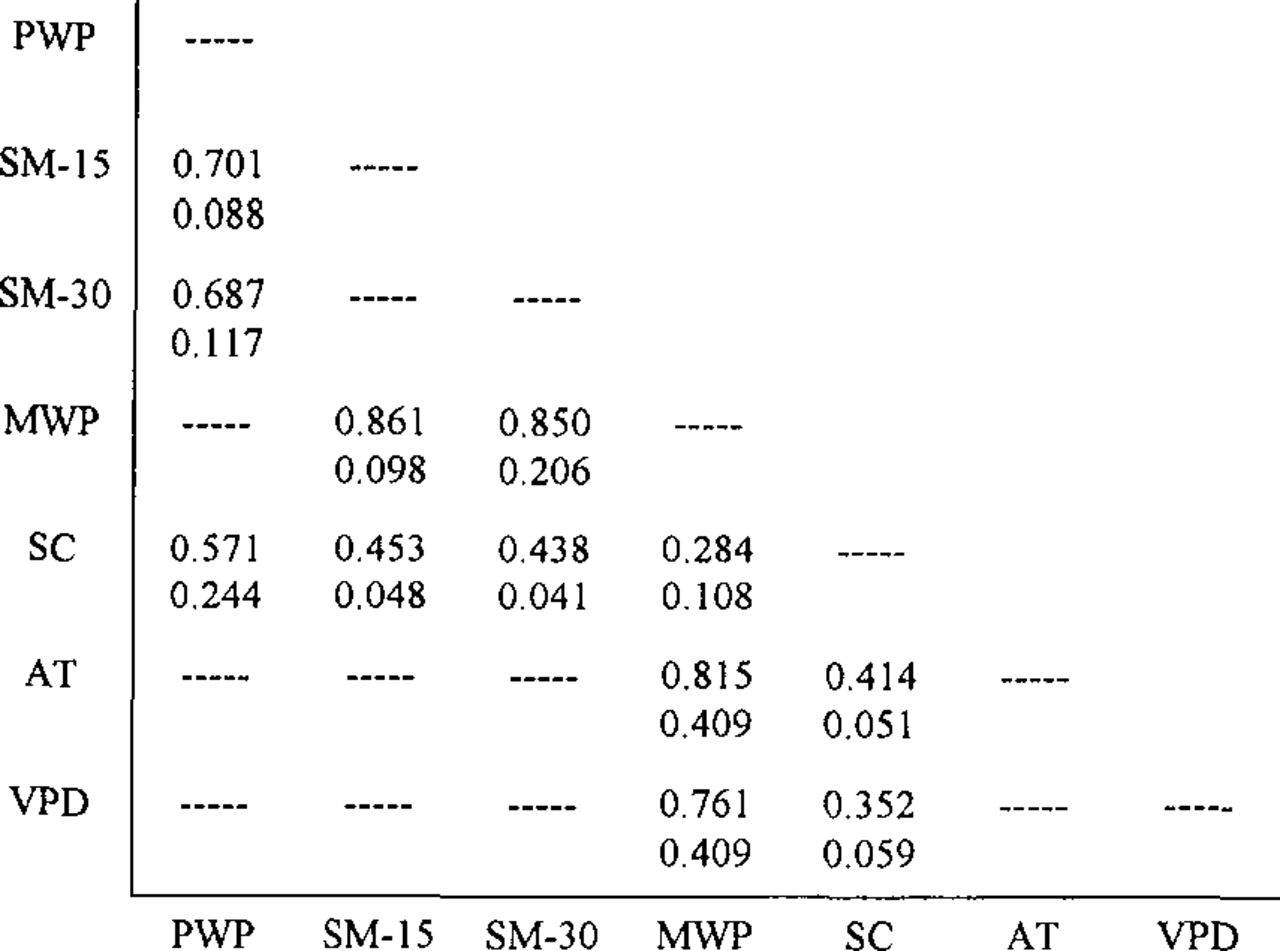
Table 1 summarizes the results of comparisons of predawn water potential, midday water potential, stomatal conductance, transpiration, and osmotic potential between the street trees (campus site) and forest trees (Minnis site). Figure 1 lists correlation coefficients relating environmental factors to water relations variables for the 2 sites. Insights can be gained and inferences made from the water relations data through comparisons between the street trees and the forest trees, and from the literature. Predawn water potential is an indirect indicator of soil moisture status and the degree of water stress being experienced by the tree (1,5,15). Predawn water potential of the street trees was significantly lower (p < 0.01) on all measurement dates, except 9/24 (Table 1). At the forest site, predawn water potential remained in a narrow range throughout the season, while values at the street site varied considerably, with a marked increase in September as higher soil moisture and lower temperatures prevailed (3). High correlations were found between predawn water potential and soil moisture at both measured soil depths of 15 and 30 cm for the street trees (Figure 1).
Tree water relations factors recorded on 6 measurement dates from July through September 1993 for street trees (campus site) and forest trees (Minnis site).

Correlation coefficients relating environmental factors to water relation variables. (PWP = predawn water potential; MWP = midday water potential; SC = stomatal conductance; SM-15 = soil moisture at 15 cm; SM-30 = soil moisture at 30 cm; AT = air temperature; VPD = vapor pressure deficit. The top numbers represent the campus site, while the bottom numbers represent the forest site.)
The forest site data were found to be in agreement with those reported by other authors for sugar maple in well-watered conditions (1,5,6,10). Predawn water potential remained at about -0.20 MPa throughout the season (Table 1). The street trees, on the contrary, were under at least a mild water stress caused by low soil moisture, with predawn water potential averaging -0.50 MPa during the summer measurements (3). The high correlation between predawn water potential and soil moisture (r = 0.70) for the street trees was consistent with other reported results (1,7,10).
Other investigators have indicated that soil moisture is not limited in urban conditions, where seasonal decreases in predawn water potential were not observed (8,15). A New York City study found excess soil moisture to be the cause of death of street trees (2). Results of the current study differ from the others, thereby pointing out the great site variability that can exist in urban conditions. There were significant differences in midday water potential between trees on the 2 sites on 4 of the 6 dates, but there was no trend (Table 1). However, for the street trees, midday water potential tended to vary with soil moisture levels and air temperature, as high correlations were found between midday water potential and each of the environmental factors on the campus site (Figure 1). Soil moisture had the strongest correlation with midday water potential (r = 0.86).
Osmotic potential results show significant differences on all measurement dates, except 9/24 (Table 1). There were no data for 9/4 due to loss of plant sample material. For the other measurement dates, the osmotic potential recorded from the street trees was significantly lower than the forest trees.
Stomatal conductance rates of Minnis forest trees were significantly higher (p < 0.01) than the campus street trees on all dates, and transpiration rates were also consistently higher on these dates. Stomatal conductance rates at the Minnis forest ranged from 122 to 170 mmol/m2/s for the 6 measurement dates, which agree with reports in the literature for well-watered sugar maples (1). The street trees, on the contrary, showed consistently low values of Stomatal conductance, and on 2 dates, the values were in the range of those found by other researchers in drought-stressed sugar maples (10).
Analysis of the street tree Stomatal conductance data showed high correlations with each of the environmental factors. Predawn water potential, as an indicator of soil moisture levels, showed the strongest correlation (r = 0.57). This relationship is consistent with results reported by Ellsworth and Reich (5), who found that predawn water potential and vapor pressure deficit were the 2 most important factors in explaining Stomatal conductance variation in sugar maple.
Employing the results of this study, along with the knowledge derived from the companion site characterization study, enables a better understanding of the water relations of the campus street trees and their adaptation to the urban site.
Relatively low predawn water potential readings are an indicator of low soil moisture availability to the roots, and a degree of tree water stress. This confirms the results of the companion site profile study, which found that the street site soil moisture levels throughout the summer were low and approached the permanent wilting point on several dates, despite the fact that rainfall was 33% above normal for the 1993 growing season. Although the midday water potential of the street and forest trees was generally equivalent during the summer measurement dates, the street trees experienced partial Stomatal closure as a means of water conservation, as noted by the consistently lower Stomatal conductance values. These results could be partially explained by the high levels of vapor pressure deficit on the street site, presented in Part I, which averaged 2.6 KPa as compared to 2.0 KPa for the forest site (3). The strongest Stomatal conductance correlations were with predawn water potential, which suggests a correlation with soil moisture levels (Figure 1). Soil moisture content could have reached critical levels, causing the roots to form abscisic acid which, when transported to the leaves, caused partial Stomatal closure. Indeed, the lowest Stomatal conductance values were associated with the periods of lowest soil moisture percentages.
In addition, these results may be further explained by possible prestressing of these sugar maples early in the season. May was a dry month, with only one-half of the normal rainfall and, as found by others (10,11,12), trees that have undergone previous water stress events develop a drought avoidance mechanism by limiting Stomatal opening in response to daily decreasing water potentials. Therefore, it could be suggested that the sugar maple street trees in this study adapted early in the season to water stress that was reinforced throughout the summer by continuing low soil moisture levels. This resulted in the reduced Stomatal conductance readings throughout the study period.
Stomatal closure would also attenuate the daily decrease in midday water potential caused by the high midday atmospheric demand that was measured in the site profile study. This could explain why the campus street tree midday water potential levels were not consistently lower than the Minnis forest trees despite the lower soil water supply and higher atmospheric demand of the street site.
Chronically low Stomatal conductances can have a serious effect on tree vitality, in that there will be significant limitations in photosynthesis due to greatly reduced diffusion of CO2 through the stomata. This can result in reduced food production for maintenance and growth. Without adequate amounts of carbohydrates produced throughout the season, growth rates are impaired as the site profile results showed (3). Also, sufficient reserves may not be stored, predisposing the tree to further environmental or biological stresses.
Osmotic potential data from this study indicate an adjustment by the campus street trees to greater water stress compared to the forest trees. This adjustment could occur by actively increasing the solute concentrations in the leaf cells, which lowers the osmotic potential, as found by others (5). Osmotic adjustment can also be passive, in that as the leaf loses water through transpiration, the relative concentration of solutes in the cells increases, thereby causing a decrease in osmotic potential. The significantly lower osmotic potentials of the street tree leaves could indicate that their turgor is greatly reduced due to greater cell water losses and the inability of the tree roots to adequately replace the lost water.
These results have provided a quantitative description on a physiologic level of how sugar maple water relations are influenced by some relatively “typical” urban site conditions, as detailed in the companion study (3). This confirms that when a mesic species such as sugar maple is placed in xeric-like urban conditions, it is subsequent water deficits that lead to its low vitality, poor growth rates, and decline.
Summary and Conclusion
The objective of any remedial actions to reduce stress in street-grown sugar maple trees would be to increase water uptake through increased soil moisture supplies and/or by increasing root volume. Evaporative demand on the site by microclimatic conditions cannot be controlled, but cultural procedures may be implemented to increase water uptake by the trees.
Supplemental watering would be a possible solution. However, at least in the case of the 210 Michigan State University street tree sugar maples that were the objects of this study, manpower and equipment costs would prohibit this solution. Additionally, permanent irrigation systems are usually not employed in the landscape, and especially in streetside tree plantings in the eastern and midwestern United States, since rainfall amounts are normally considered sufficient for tree growth.
Removal of the turfgrass from within the dripline of the trees and replacement with organic mulch would greatly increase fine root density as well as improve soil moisture levels (14). Two additional treatments appear to hold promise for the urban soil conditions encountered in this study, namely, radial trenching, which has been studied at Cornell University (4), and preliminary results of research by Kielbaso at Michigan State University with the “Grow Gun” (unpublished). The purpose of treatments such as these is to create a better soil environment for root growth and development, which would eventually lead to increased water uptake to meet the demands of the tree crown.
In terms of selecting trees for new planting, this study reiterates the sensitivity of sugar maple to water deficits. However, work done by Pair (9) has demonstrated that certain cultivars of sugar maple (e.g., Caddo and Legacy) show a greater tolerance to drought stress. These sugar maple cultivars should be considered by urban foresters in the future for street site plantings.
This water relations study and its companion site characterization profile have focused on an urban street site that might be considered less harsh than many street sites surrounded by more paving and buildings. Similar studies with other tree species in varying urban conditions would provide needed quantified descriptions of the urban ecosystem and the physiologic response of tree species growing in these ecosystems. With more of this type of information, arborists and urban foresters will be able to make better decisions regarding treatments for stressed trees. In addition, they will be in a better position to provide improved species and cultivar selections, and recommend landscape specifications for streetsite plantings.
Acknowledgments
The authors wish to acknowledge the efforts of the following persons from Michigan State University in the design and execution of this research: Dr. Carl Ramm, Dr. James Flore, Mr. Andrew Burton, Mr. Paul Swartz, Ms. Peggy Payne, Mr. Andrew David, and Ms. Sharon Bahn. Critical comments of reviewers on an earlier draft of the manuscript are truly helpful and appreciated.
Footnotes
- © 1996, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.