
Abstract
In 1982 Carter reported two mechanisms in the resistance of certain species of lindens (Tilia spp.) to the linden aphid, Eucallipterus tiliae: simple pubescence and glandular prominences on the underside of the leaves. Field studies conducted from 1991-93 confirmed that aphids reach higher numbers on linden species with reduced pubescence (T. cordata and T. europea) than on a more pubescent species (T. platyphyllos). The rate of aphids parasitized by the introduced parasitoid Trioxys curvicaudus is also higher on the latter species.
The linden aphid, Eucallipterus tiliae, feeds only on lindens (Tilia spp., known as lime trees in Europe) (5). It was probably introduced to North America from Europe in the late 1800’s, and is now wide-spread across the United States and Canada (6,7). A biological control program in 1970 resulted in the establishment of Trioxys curvicaudus in northern California; initial studies reported good control of the aphid at the release sites (4,8). However, research conducted by our lab in the 1980’s indicated that the aphid was not under control at other sites, prompting our present studies into the aphid and its natural enemies.
Some linden species appear naturally resistant to the aphid in Europe; in Britain, Carter (2) reported that resistance can be attributed to two mechanisms. On T. petiolaris, the pubescence on the underside of the leaf interfered with feeding by young aphids. In three other species (T. maximowicziana, T. euchlora, and T. mongolica) resistance was attributed to the presence of “small glandular prominences” at the aphid feeding sites. Those species which Carter considered lacking resistance included small-leaved lindens (T. cordata), large-leaved lindens (T. platyphyllos) and a hybrid of these two, the common linden (T. europea).
From 1991-93 we studied the linden aphid and its natural enemies in northern California on three linden species. This paper reports our findings on the species effect on aphid numbers and rates of parasitism by three Trioxys species, T. curvicaudus, T. pallidus and T. tenuicaudus (the latter two species were established in this area in biological control programs against the walnut aphid in 1968, and the elm aphid in 1972, respectively).
Methods
We sampled at weekly intervals from June-November in 1991 and May-October in 1992, and at biweekly intervals from May-November in 1993. Eight sites in five counties were chosen: three in Alameda Co. (two in Berkeley, one in Pleasanton), two in Marin Co. (one each in San Anselmo and San Rafael), and one each in Napa Co. (Napa), Santa Clara Co. (San Jose) and Solano Co. (Vallejo). Four sites were planted with T. platyphyllos, three with T. cordata, and one with T. europea - species determinations were made using Bailey (1) (cultivar or subspecific determinations were not made). All trees were fairly well-established; their ages were unknown, but ranged in height from 6-7 m (16 cm dbh) to about 18 m (90 cm dbh).
Trioxys curvicaudus was present at all sites in Alameda and Santa Clara County, and initially absent from the other three counties. In 1992 we collected parasitized aphids from Berkeley and San Jose; emerging T. curvicaudus females and males were held together and observed for signs of mating. From 1 June to 26 October we released 77 mated and 32 unmated females and 12 males at the San Rafael site. Recoveries from parasitized aphids were made in San Rafael in late 1992 and 1993, indicating successful establishment.
Three or four trees were sampled at each site; on each sampling date, eight samples were taken from the lower canopy of each tree (that part accessible from an 8-foot ladder), one each from the inner and outer canopies of the four cardinal directions (N,E,S,W). Each sample consisted of all the leaves on the terminal one foot (30 cm) length of a branch. Both aphids and natural enemies were counted in the field, and aphids were collected and dissected in the lab to calculate the percentage parasitism. Field counts were tabulated as the mean number aphids/sample/tree/site/date and transformed using log (x+1) conversion to eliminate non-homogeneous variances; parasitism was tabulated as the mean percent parasitism/site/date. Since some sites were sampled more often than other sites, we choose to analyze only those dates when all 8 sites were sampled (10 in 1991, 14 in 1992 and 10 in 1993).
Results
Values from the 34 dates are averaged for each site and presented in Table 1 (San Rafael is split by years to delineate the effect of T. curvicaudus after it was introduced in 1992). The trees at the San Anselmo site appeared lacking in vigor - they were the smallest T. platyphyllos noted in our study, with a large proportion of dead branches. This stand consistently had extremely low numbers of aphids and natural enemies, and we believe some unidentified systematic factor was operating there which depressed the aphid population. Therefore this site was excluded from further analysis.
We analyzed aphid numbers with a 2-way ANOVA test (using tree species and the presence/absence of T. curvicaudus as categories). The effect of tree species was highly significant (p<.0005); T. europea had the highest populations (by 1-2 orders of magnitude), T. cordata intermediate populations, and T. platyphyllos the lowest. Although there was no significant difference in aphid numbers at the T. europea site before and after T. curvicaudus was established, the presence of T curvicaudus was highly significant (p=.009) overall, while the interaction between tree species and T. curvicaudus presence wasn’t significant at the .05 level.
Trioxys curvicaudus was the dominant parasitoid in Berkeley, Pleasanton and San Jose, accounting for 90% of all reared Trioxys species; T. pallidus represented 2% and T. tenuicaudus 8%. At the other sites, T. pallidus was dominant, representing 100% of the Trioxys spp. The only exception was at San Rafael, where T. curvicaudus accounted for 0% of the Trioxys spp. in 1991 and May-June 1992, 65% from July-Nov. 1992, and 82% throughout 1993. The parasitism rates at the sites where T. curvicaudus was present varied inversely proportional to aphid numbers. A 1-way ANOVA test showed parasitism was significantly higher on T. platyphyllos (about 20%), than on T. cordata (about 5-7%), or T. europea (about 2%) (p<.0005).
Linden aphid populations and Trioxys spp. parasitism from 8 northern California sites, 1991-93.
Discussion
Two factors limiting linden aphid populations appear to be operating in northern California: the linden species planted, and the presence of T. curvicaudus. These factors are additive, so aphid numbers are lowest and parasitism rates highest in stands of T. platyphylloswhere the parasitoid is present; aphid numbers are highest on T. europea and intermediate on T. cordata.
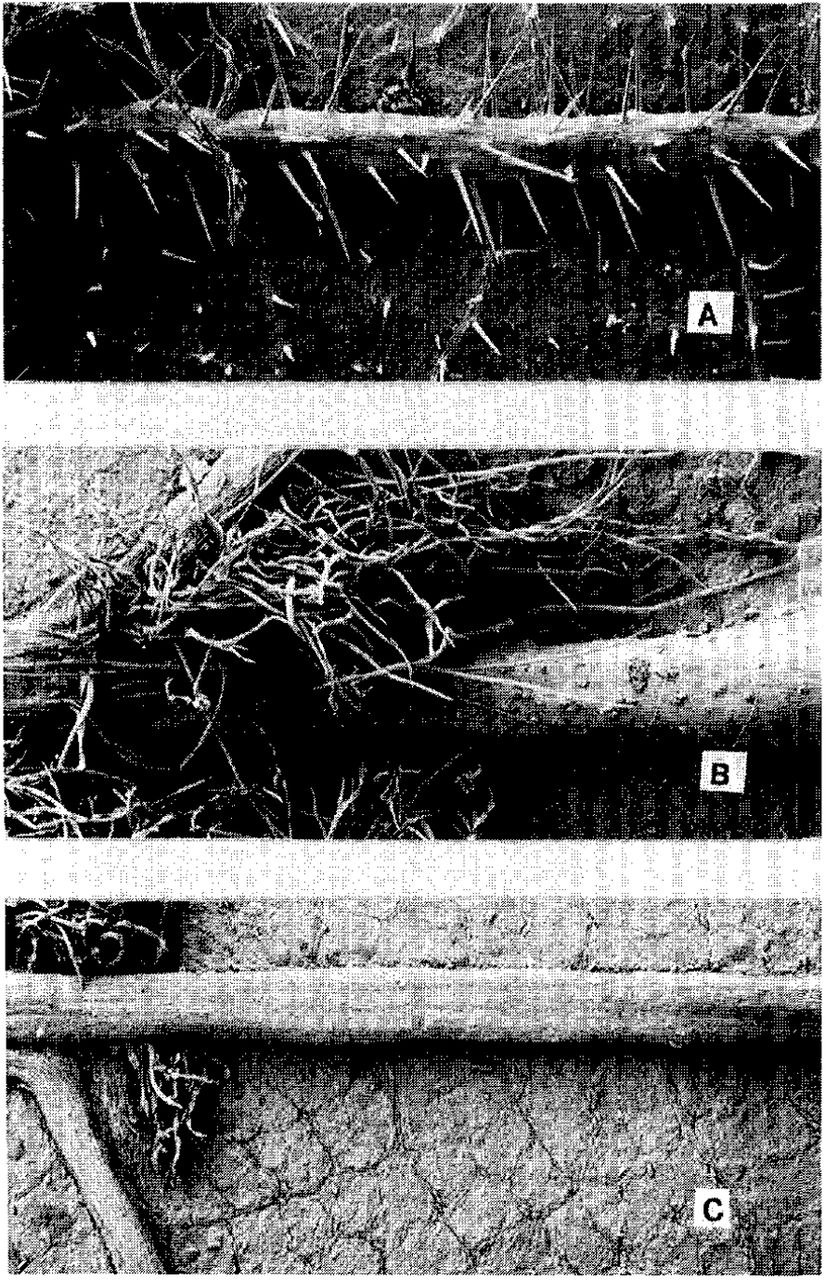
This situation is consistent with the mechanism of resistance due to leaf pubescence discussed by Carter (2): T. platyphyllos has numerous hairs around the leaf veins, which could interfere with the ability of younger aphids to feed at the favored sites near the veins, while the hairs of T. cordata are fewer and more recumbent, and those of T. europea are restricted to the axillary tufts (Fig. 1).

Scanning electron micrographs of midvein on ventral leaf surface. A - Tilia platyphyllos, with erect pubescence concentrated on veins and scattered elsewhere (x 35.7); B - T. cordata, showing axillary tufts and reduced, recumbent pubescence on midvein (x 36.7); C - T. europea, showing lack of pubescence, except for axillary tufts (x 35.9)
Interestingly, Carter considered T. platyphyllos susceptible to the aphid. However his observations may have been made in the absence of T. curvicaudus, which is known only from the southern part of Great Britain. A similar condition exists at Napa (where only T. pallidus is present), where aphid numbers were higher and parasitism rates lower than in Berkeley and San Jose (where T. curvicaudus has been established for 15-20 years).
The lower aphid numbers and higher parasitism rates at sites where T. curvicaudus has been long-established (Berkeley, Pleasanton and San Jose) demonstrate that this species can be an important mortality agent of the linden aphid. However, this parasitoid did not reduce the aphid population in San Rafael, for which we offer two possible explanations. First, the aphid numbers resulting from the lack of resistance in T. europea may be too high for T. curvicaudus to overcome. Second, the parasitoid was only introduced there in 1992, and requires more time to build up its numbers to be effective.
Since the linden aphid is well-established throughout the continent, arborists should consider planting only those linden species which show some degree of resistance. In the absence of T. curvicaudus, T. platyphyllos should be considered a susceptible species. Care must also be exercised due to varietal differences - eg.: T. platyphyllos subspecies pseudorubra has virtually hairless leaves (3).
Acknowledgments
We thank Joe Bates of Vallejo, Mark Beaudoin of San Jose, Frankie Campbell of San Rafael, Robert Carlsen of Napa, William Jones of Berkeley, Larry Messa of Pleasanton and Dean Nyberg of San Anselmo for their cooperation in allowing us access to the sites. David Rowney of the Laboratory of Biological Control, University of California, Berkeley provided help with the statistical analyses. Thanks also to Wilfred Bentham of the Robert D. Ogg Electron Microscope Laboratory, U.C. Berkeley, for preparing and processing the micrographs, and to an anonymous reviewer for providing helpful suggestions for the paper.
- © 1994, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.