
Abstract
This report summarizes the findings of four augmentative releases of insect predators, lady beetles and lacewings, for controlling aphids on landscape plants. Releases of the convergent lady beetle and seven spotted lady beetle reduced populations of spirea aphid on firethorn when dispersal was limited. Neither species of lady beetle reduced populations of woolly aphids on firethorn. A field release of convergent lady beetles was followed by reduced aphid populations on Elaeagnus in a landscape. Releases of lacewings on stonecrop and hawthorn provided no evidence of reductions in aphid populations. Some impediments to the success of augmentative releases in landscapes are discussed.
Several types of pest control are available to arborists, landscapers, and producers of landscape plants to help reduce damage from insects and mites. Traditionally, there has been heavy reliance on chemical pesticides. In recent years this approach has become less attractive, prompting increased interest in the development and implementation of alternative approaches. From an ecological standpoint it is clear that when insecticides are used unwisely problems can arise. These problems include development of pesticide resistance, pest stimulation, pesticide-induced outbreaks, damage to non-target organisms, and adverse effects on human health. The ecological impact of pesticide use in landscape systems has been discussed at length elsewhere (18,19).
In addition to ecological concerns regarding pesticide use there are now mounting and interrelated sociological, economic, and regulatory incentives for considering alternatives to chemical controls. Perhaps foremost among these incentives is consumer demand. Pesticide residues around homes and public institutions are less acceptable to the public than in the past, and landscape managers consequently find themselves confronted with requests for nonchemical controls. Chemical pesticides are also being subjected to increasingly restrictive government regulation.
Government regulation may affect management of landscape plants in two ways. First, the cost of registration and re-registration of agricultural chemicals can be prohibitive. As a result, some chemicals become unavailable for landscape use requiring the use of alternative chemicals or alternative management strategies. Second, new regulations are being aimed directly at the use of pesticides in landscapes. At least a dozen states currently have enacted legislation requiring the posting of signs in areas treated with pesticides. Several states require written notification to neighbors or pesticide-sensitive individuals adjacent to application sites (1). This increased regulation represents a trend which may be expected to continue as increased public awareness and concern affects governmental policy.
One alternative to the use of insecticides for insect and mite control on trees and shrubs is the use of biological control. Biological control is the use of predators, parasites, and pathogens to reduce pest populations below damaging levels (21). Biological control can be implemented or enhanced in landscapes in a variety of ways, however, there are four general approaches; conservation, importation, formulation and augmentation (6,10,18,19,21).
Conservation means that existing natural enemies are protected through management practices that favor survival and reproduction. This includes altering timing, methods of application of materials, or using non-chemical control tactics such as pruning or mulching (18,19). In importation, foreign natural enemies are introduced to control a pest. Often the pest itself is a foreign invader, but attempts have been made to import natural enemies for the control of native pests. This approach has been used with dramatic success to control landscape pests such as the ash whitefly and eucalyptus longhorned borer in California (16). A third approach to biological control includes the formulation of pathogens or nematodes which can be applied in a manner similar to chemical pesticides. The use of the bacterium, Bacillus thuringiensis (Bt), and entomopathogenic nematodes for control of foliar feeding and boring caterpillars has been the topic of recent articles in this journal (4,5,9). Finally, when existing natural enemies are too scarce or appear too late to prevent damage, extra parasitoids or predators, purchased from a commercial insectary orcollected elsewhere, can be released. This approach is called augmentation.
Augmentative releases have proven highly effective in a variety of crop production systems, including vegetables, fruits, greenhouses, and interiorscapes (10,11,12,20). This approach has recently generated much interest for controlling insect and mite pests in landscapes and gardens (7) and at least two recent surveys indicate that arborists are using releases of predators and parasitoids for pest control on a limited scale (8,15). However, there are few published accounts evaluating the efficacy of augmentative releases in landscape settings.
The purpose of the studies reported here was to evaluate the efficacy of two predator groups, lady beetles (Coccinellidae) and lacewings (Chrysopidae) for reducing populations of aphids on landscape plants. These organisms were chosen for two reasons; aphids are key pests of trees and shrubs throughout the United States (22), and both groups of predators are readily available from commercial suppliers and are promoted for aphid control in landscapes. Two of the releases reported here were controlled, experimental studies designed to specifically determine the efficacy of lady beetles and lacewing releases against aphids on firethorn and stonecrop. We also report the results of two augmentative field releases, lady beetles on Elaeagnus infested with aphids, and lacewings on hawthorn infested with aphids. The authors recognize that the results of the latter two releases must be interpreted with caution. They are of interest because they represent practices of landscape managers, and because of the dearth of information on augmentative releases of natural enemies in the landscape.
Materials and Methods
Lady beetles for control of aphids - greenhouse studies
The first series of experiments evaluated the use of two species of lady beetles for reducing aphid populations on the woody landscape shrub, firethorn (Pyracantha lelandii). Sixty firethorns in three gallon containers were placed in a vented greenhouse and infested with two species of aphids. Plants averaged 16.3 (s.e. = 0.3) inches in height and 12.8 (s.e. = 0.3) inches in spread. Aphids used in the study were the spirea aphid, Aphis spiraecola, and a woolly aphid, Eriosoma sp. Both species of aphids are common pests of rosaceous plants including apple, hawthorn, and firethorn as well as several species of landscape plants (13,14).
Two common species of lady beetles were used for the study. Hippodamia convergens, the convergent lady beetle, is a native coccinellid and widely available from commercial vendors. Coccinella septempunctata, the seven spotted lady beetle, is an imported species that is widely distributed throughout much of the United States (2). Both feed on a wide variety of aphids and other soft bodied insect prey (11). Convergent lady beetles used in this study were purchased from a commercial vendor of biological control agents and seven spotted lady beetles were obtained from naturally occurring field populations.
Populations of both aphids were allowed to develop for approximately two months prior to the release of lady beetles. When the plants were heavily infested, they were randomly divided into two groups of thirty. The first thirty plants were then randomly assigned to one of three treatments. Ten plants were placed in cages made of window screening to allow aphid populations to develop in the absence of lady beetle predation. The next ten plants also were enclosed in cages and ten convergent lady beetles were added to each cage to estimate the effect of the predator when migration of the beetle was prevented. The final ten plants were uncaged for the duration of the study and each received ten lady beetles which were allowed to migrate.
Immediately prior to the release of lady beetles all plants were misted. Releases were made at 5:30 p.m. at an ambient temperature of approximately 85° F. At the time of predator release and at two (day 2) and seven days (day 7) after the release, all aphids were counted and identified to species on four shoots of each plant. At two and seven days after the release the H. convergenson each plant were counted. The remaining thirty plants were similarly randomly assigned to the same three treatments, with C. septempuntata released onto the plants. The abundance of aphids and seven spotted lady beetles was observed at the same time intervals as in the previous study. At each observation the numbers of aphids and lady beetles were compared among the three treatments using an appropriate test statistic. Prior to the analysis, variances of the mean number of aphids found per shoot or total number of lady beetles found per plant were compared with a Bartlett’s test for homogeneity (23). When th is test satisfied the assumption of homogeneity, the experiment was analyzed using an analysis of variance (ANOVA) for a completely randomized design (23). When the assumption of homogeneity was not met, a Kruskal-Wallis (KW), nonparametric analysis was used (23).
Lady beetles for control of aphids - field release
The plants used in the study were Elaeagnuspungens ‘fruitlandii’ planted in a hedge that was approximately 48 inches tall, 48 inches wide and 108 inches long. The shrubs were heavily infested with the aphid, Capitophorus elaeagni, a species known to infest thistles and artichokes in addition to Elaeagnus (17). Convergent lady beetles were purchased from a commercial vendor, hydrated in their shipping container with a spray of water, held in a refrigerator overnight at 45° F, and released at 8:00 a.m. onto dew moistened Elaeagnus when ambient temperatures were 72° F. Fifteen hundred lady beetles were distributed uniformly throughout the hedge by releasing the beetles at the bases of several plants.
The effect of the lady beetles on aphids was evaluated by comparing aphid abundance at the time of the release with their abundance 15 days after the release. At both time intervals aphid abundance was estimated in two ways. First, 100 randomly selected shoots were examined for the presence or absence of aphids; then, on 10 of the shoots bearing aphids, the number of aphids was counted on each leaf and the average number of aphids per leaf calculated. The frequency of infested shoots was compared between the release date and 15 days post release with a Chi-square test for heterogeneity. The average number of aphids per leaf was compared between dates with a Kruskal-Wallis test.
In addition to recording aphid abundance, the abundance of convergent lady beetles was also observed. Prior to the release of beetles, five minute counts of convergent lady beetles were made at three different locations in the canopy of the plants. This procedure was repeated 15 days after the release.
Lacewings for control of aphids on stonecrop and hawthorn
The first experiment evaluated the ability of the lacewing, Chrysoperla rufilabris, to reduce aphid populations in a landscape setting. The plant used in the study was an herbaceous perennial stonecrop, Sedum spectabile. The trial was conducted in six raised planter boxes, each of which contained 4 stonecrop plants. We documented that the stonecrop plants were slightly smaller on average where the lacewings were released compared to control plants. Release plants averaged 1,796 (s.e. = 1,081) cu. inches and control plants averaged 4,943 (s.e. = 3,171) cu. inches. There was one Chinese elm tree in the center of all but one box. The boxes were not under irrigation. The stonecrop plants were infested with Aphis sedi.
Three planter boxes were randomly selected to receive releases of lacewings and three boxes were randomly assigned to be controls. For the treatment, approximately 55 lacewing larvae were released on each plant in the box (220/planter box). No lacewings were released in boxes assigned to the control treatment.
Chrysoperla rufilabris larvae were obtained from two commercial vendors. Lacewings were packaged as eggs in corrugated cardboard cells covered on both sides with organdy. The larvae from the first vendor were shipped from a producer to the vendor and then to our laboratory where they arrived approximately four days after they left the producer. They were received in a styrofoam cooler, but without any cooling source. An unacceptable lacewing mortality of approximately 67% occurred in the packages as supplied.
Lacewings ordered directly from the producer were shipped in a cooler with ice packs, and arrived within approximately 24 hours of shipment. The larvae were stored overnight in the shipping cooler with the ice packs and some damp newspaper. At 6:00 a.m. on the day after arrival, counts were made of live lacewing larvae in five, 10-cell replicates. The average number of larvae per cell was calculated from these counts and used to estimate survival during shipping and storage and to estimate the number of lacewing larvae released per plant. The vendor claimed that approximately 1 larva per cell would be received, and an average of 1.1 live larvae per cell were counted on the morning of release, indicating satisfactory survival during shipment and storage.
At the time of release, the cardboard packaging was cut into 50-cell blocks (each block containing approximately 55 larvae), and the excess larvae (from cut cells) were shaken off the packaging away from planter boxes. The blocks were then held over plants and the organdy was removed. Lacewings were gently shaken or brushed off the organdy onto the plants. After the cells were opened, the block was placed in the interior of the plant to allow lacewings to crawl from the packaging onto the plant. Releases began at 7:30 a.m. and were completed by 7:40 a.m. The ambient temperature at the time of the release was approximately 70° F.
At the time of release the abundance of aphids was estimated by counting the aphids on two randomly selected leaves on each plant in each box. During counts, the presence or absence of ants in planter boxes was noted. Counts of aphids were made 1 week after release, by the same method used at the time of release. The presence of ants in boxes was noted again. The average number of aphids per leaf was compared for plants with and without lacewings before and seven days after release using ANOVA or KW. Nine days after release, the stonecrop and elm trees in planter boxes were checked after dark (9 p.m.) for lacewing larvae. Because C. rufilabris is reported to be nocturnal (6), the survey on day nine was made at night. The elm trees in the planter boxes were checked as well as the stonecrop.
A second experiment attempted to use commercially supplied lacewings to control populations of bean aphid, Aphis fabae, infesting Washington hawthorns, Crateagus phaenopyrum, in a landscape setting. Bean aphid is one of the most common aphids found attacking a wide variety of plants in the landscape (13). Twelve trees approximately 8 ft tall were heavily infested with the aphids.
The lacewings used in the study were a combination of Chrysoperla carnea (common green lacewing) and C. rufilabris, packaged as hatching eggs attached to paper bands. One band of lacewing eggs was attached to the bole of each hawthorn. This procedure delivered 200-300 lacewings eggs per tree. The eggs were placed on the trees at 8:00 a.m. and the ambient temperature at the time of release was 75° F.
On the release dates, aphids were counted on two randomly selected shoots on each tree and an average was computed. Fifteen days after the release of lacewings two randomly selected shoots were examined again for aphid abundance. In addition to examining shoots for aphids, the canopy of each tree was examined for five minutes and the number of lacewings recorded. The average number of aphids present on shoots fifteen days after the release was compared to the number of aphids present at the time of release with ANOVA.
Results
Lady beetles for control of aphids - greenhouse studies
Both species of lady beetles appeared to reduce spirea aphid numbers when released in cages. At the time of release of H. convergens abundance of spirea aphids did not differ among treatments (ANOVA, F = 2.82, P = 0.09). However, two and seven days after the release of H. convergens, aphid abundance differed among treatments (day 2, KW, H = 8.34, P = 0.015; day 7, KW H = 10.03, P = 0.007). The trend clearly was for aphid populations to be lower on plants where lady beetles were enclosed in cages (Figure 1). Uncaged plants and caged plants with no beetles exhibited no differences in aphid populations. The same pattern was observed for releases of C. septempunctata (Figure 1). At the time of release, aphid numbers differed somewhat between treatments (ANOVA, F = 4.18, P = 0.04) and were slightly higher on caged plants where no lady beetles were released. On days 2 and 7, treatments differed significantly in the numbers of spirea aphids (day 2, KW, H = 6.23, P = 0.043; day 7, KW, H = 9.43, P = 0.009). As was the case with H. convergens, spirea aphids were much less abundant on plants enclosed in cages where lady beetles had been released.
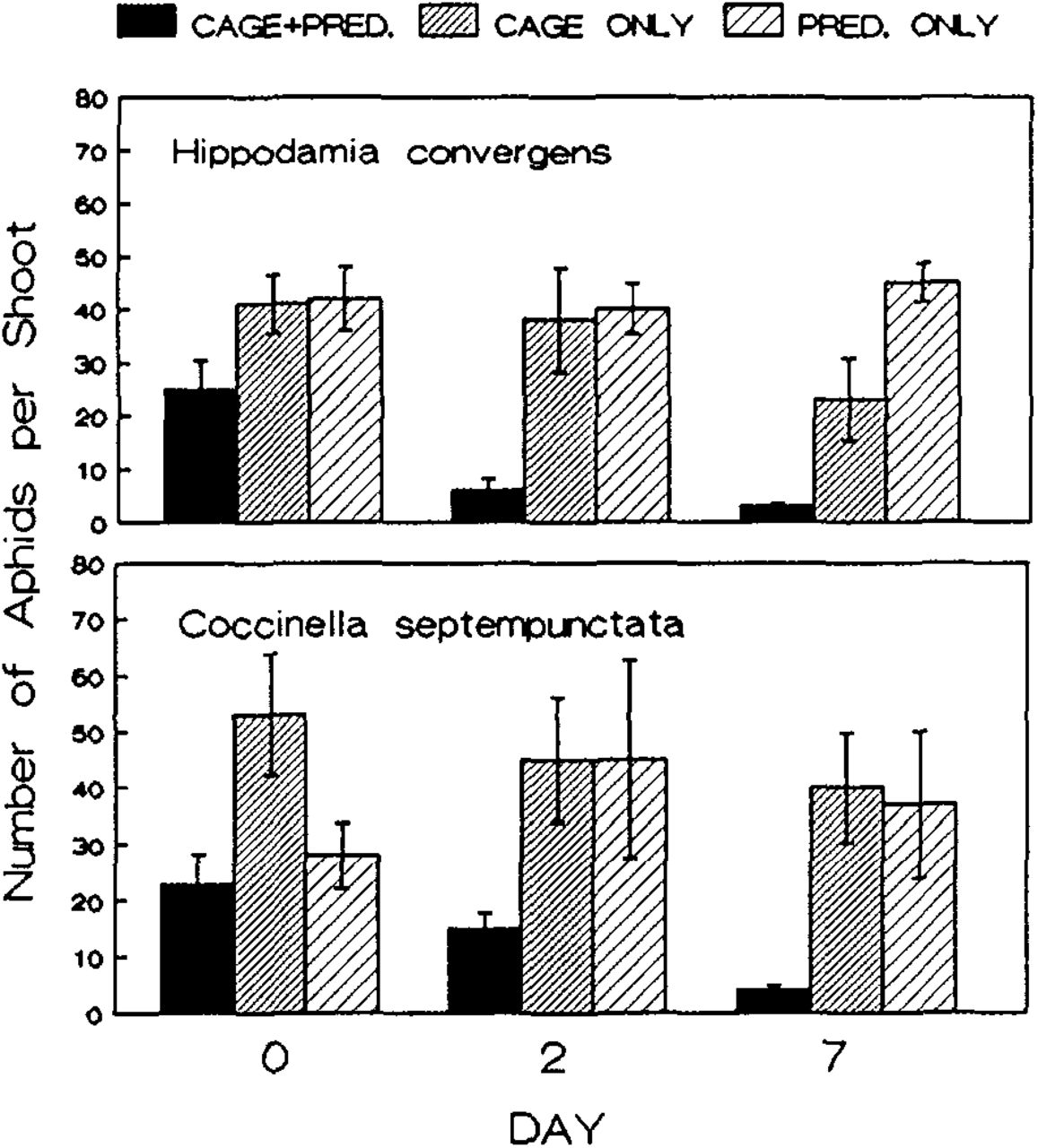
Effect of Hippodamia convergens and Coccinella septempunctata on the abundance of Aphis spiraecola on caged and uncaged firethorns. Bars represent means and vertical lines represent standard errors.
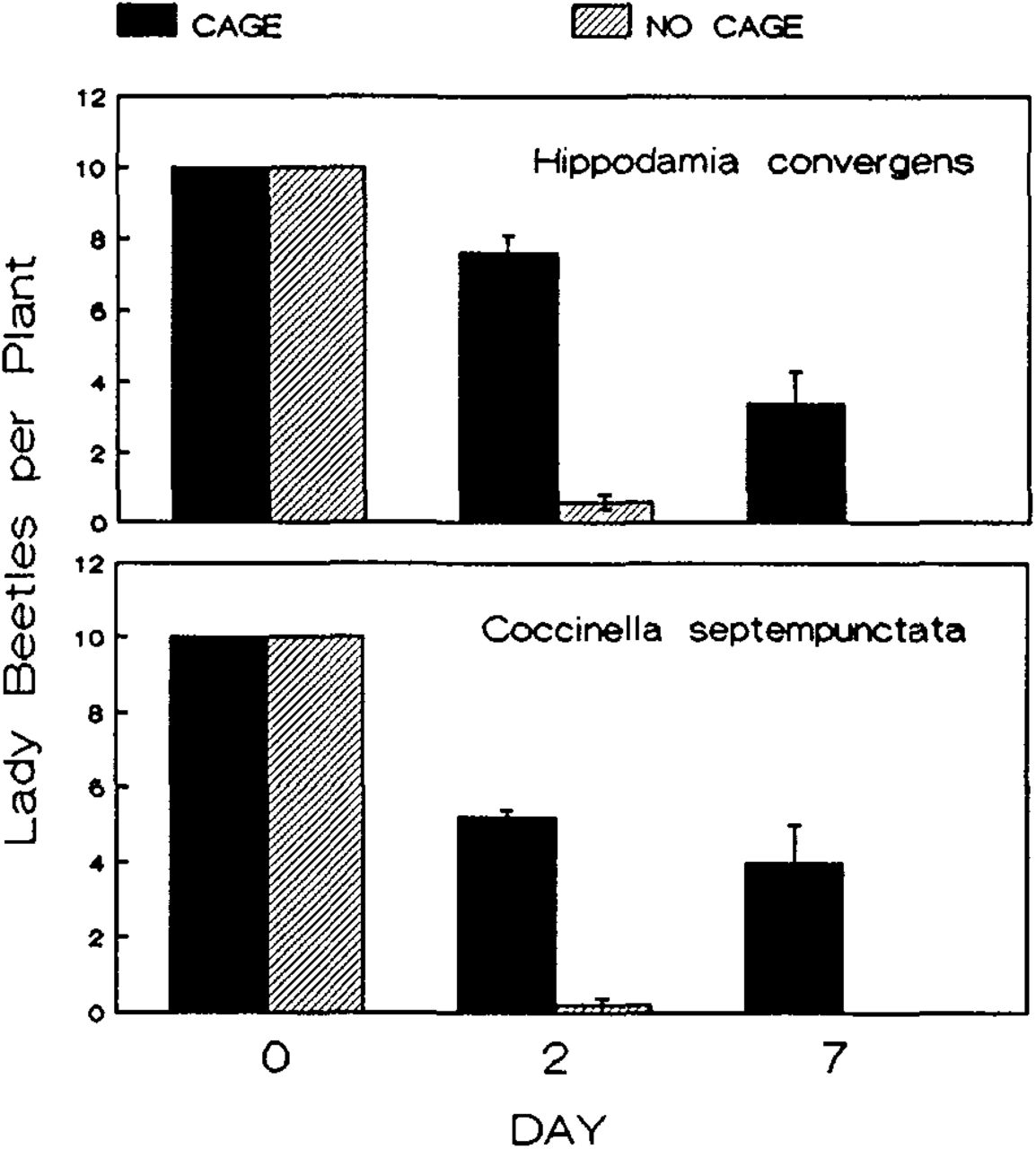
An examination of lady beetle abundance on plants provides insight into why aphid abundance varied among treatments. Both species of beetles rapidly dispersed from uncaged plants and failed to return to aphid infested plants during the study period. By day 2, uncaged plants had significantly fewer H. convergens (ANOVA, F = 153.13, P = 0.000), and C. septempuntata (KW, H = 6.73, P = 0.009) than caged plants (Figure 2). This pattern held for the remainder of the study. The only treatment where beetles were abundant and reduced spirea aphid populations was where plants were enclosed in cages.

Changes in the abundance of Hippodamia convergens and Coccinella septempunctata and following their release on caged and uncaged firethorns. Legend as in Figure 1.
Neither species of lady beetle was effective in controlling the woolly aphid on pyracantha. For H. convergens there were no differences between treatments in woolly aphid abundance on firethorns at the time of release, two days after release, or seven days after release. Likewise for C. septempunctata, woolly aphid populations did not differ significantly among treatments at the time of release or at either of the subsequent intervals of observation.
Lady beetles for control of aphids - field studies
At the time of the release beetles were observed moving immediately into the canopy of the plant. Shoots of Elaeagnus were significantly less likely to be infested with aphids fifteen days after the release of H. convergens than before its release (Chi-square = 61.44, P = 0.000). Prior to the release of beetles, 100% of the shoots were infested with aphids, and 15 days after the release only 53% of the shoots were infested. A similar trend was seen in the abundance of aphids on plant leaves. Prior to release, there was a mean of 24.03 (s.e. = 8.41) aphids per leaf, whereas 15 days after the release there were 5.5 (s.e.= 1.05) per leaf. This difference was significant (KW, H = 10.09, P = 0.000) and represents a 77% reduction in aphid abundance. Prior to release, no H. convergens were observed in the hedge in three five-minute counts, whereas a mean of 12.33 H. convergens per five minutes were observed in three five-minute counts taken on the fifteenth day after release.
Lacewings for control of aphids on stonecrop and hawthorn
Lacewing larvae did not appear to have an impact on aphid density on stonecrop plants (Figure 3). The mean aphid density at the time of release was significantly higher in control planters (KW, H = 3.86, P = 0.05). One week after the release of lacewings, there was no significant difference in aphid numbers between treatments (ANOVA, F = 0.66, P = 0.46). The change in aphid density between the release date and seven days post release also did not differ significantly between treatments (ANOVA, F = 0.02, P = 0.894).

Effect of Chrysoperla rufilabris release on the abundance of Aphis sedi on stonecrop. Legend as In Figure 1.
Lacewing larvae did not appear to persist in the planter boxes after release. No lacewing larvae were observed at the seven day post-count or during the nocturnal survey that took place nine days after release. Ants were present and associated with aphids, in all boxes except for one replicate of the control treatment.
There was no difference in aphid density on hawthorn from the date of lacewing release (mean = 129.8, s.e. = 17.8) to the sample date fifteen days later (mean = 134.8, s.e. = 26.7) (ANOVA, F = 0.02, P = 0.82). Although no control trees were available for comparison, it is clear that aphid populations, on average, did not decline following the release of lacewings. No lacewings were observed on any plants fifteen days after the release.
Discussion
Huffaker (10) suggested that augmentation is best attempted when the natural enemy is known to be effective in regulating pest populations but where conditions such as weather or asynchrony with the host prevent control from occurring. While releases of lady beetles and lacewings have proven successful in reducing aphid populations in crop, fruit, and greenhouse systems (10,11,12,20), very few studies have been undertaken to investigate their use in landscape systems. A notable exception is the work of Booth (3) who found multiple releases of the convergent lady beetle to effectively reduce populations of crepemyrtle aphid. Lacewing larvae were not effective in reducing populations of this pest (Booth, personal communication). The results reported here suggest the possibility of implementing augmentative releases in landscape and nursery settings.
First, from the releases of lady beetles three conclusions were reached. Both the convergent and seven spotted lady beetle have the potential to reduce populations of the spirea aphid if emigration can be prevented. This may be practical in a greenhouse situation where beetles are confined by a structure or in a nursery or landscape situation where small plants can be enclosed in netting. Confining predators may not be practical for arborists or landscapers involved with the management of this aphid on large or numerous plants or where the presence of netting is objectionable. Furthermore, the convergent lady beetle appears to provide substantial levels of control of the aphid, C. elaeagni, on Elaeagnus in a landscape setting.
The results pertaining to the convergent lady beetle are somewhat surprising for the following reason. DeBach (6) cautioned against the use of convergent lady beetles that had not been pre-conditioned priorto their release. Beetles collected from overwintering sites might be in a physiological state not conducive to feeding and reproduction and could exhibit a migratory behavior upon release. The supplier of convergent lady beetles used in these studies indicated that beetles had not been preconditioned prior to shipment. Nonetheless, convergent lady beetles confined to cages fed on spirea aphids and reduced populations within two days of their release. Also, many of those released into the landscape apparently colonized Elaeagnus plants, consumed aphids, and remained at the site at least fifteen days after their release. The possibility that the decline in aphids on Elaeagnus was only coincident with the release of lady beetles but caused by other factors such as changes in weather, the host plant, or phenology, however, cannot be eliminated.
The release of lady beetles on firethorns also demonstrated that when firethorns were simultaneously infested with spirea and woolly aphids, woolly aphids were not effected by either convergent or seven spotted lady beetles. This indicates that in open landscapes, where a variety of prey species are available to released natural enemies, the relative acceptability of the target pest may affect the outcome of an augmentative release.
Finally, the release of lady beetles on firethorns indicated that both species of lady beetles rapidly dispersed from firethorns heavily infested with aphids when not confined by cages. Clearly, this finding is consistent with Debach’s (6) caution to use only preconditioned H. convergens. However, the rapid migration of C. septempunctata, which were actively feeding and reproducing immediately prior to release is not consistent with this explanation. Other, yet unknown, factors prevented the successful establishment of seven spotted lady beetles on uncaged plants.
Releases of lacewings for control of aphids on stonecrop and hawthorn did not prove successful. However, we demonstrated that lacewing mortality can be unacceptably high when the interval between the harvest of lacewing eggs and the arrival of neonate larvae at the release location is extended beyond.a few days due to delays in shipping. DeBach (6) cautioned that lacewing eggs and hatchling larvae should be placed at the release site as rapidly as possible. By purchasing larvae directly from a producer, shipping time and associated mortality were minimized and we recommend this procedure.
The reasons for the failure of lacewings to control aphids on stonecrop and hawthorn are unclear. It is worth noting that lacewings did not appearto persist in the landscape in either release reported here. The presence of ants was noted in each of the planters where lacewings were released on stonecrop. DeBach (6) and Huffaker (10) suggested that ants could play an important role in limiting the efficacy of biological control agents by protecting honeydew producing pests such as aphids. It is possible that the ants observed in the planters prevented lacewings from attacking aphids on stonecrop. Furthermore, DeBach (6) indicated that lacewing larvae could rapidly dehydrate unless plants provided daytime refuge with sufficiently high levels of humidity. Although the stonecrop plants had dense canopies, it is possible that insufficient moisture was available for lacewings to persist in the planter boxes. Regardless of the underlying reasons for the lack of aphid population reductions associated with the release of lacewings, we concluded that lacewings released in the manner and at the rate attempted here would not likely provide the level of control sought by arborists, landscapers, or nursery managers. Booth (personal communication) similarly found that releases of lacewings failed to reduce populations of crepemyrtle aphids on crepemyrtle.
In summary, these studies demonstrated that under certain conditions both the convergent and seven spotted lady beetle significantly reduced populations of spirea aphid on caged firethorn. In a landscape setting there was good circumstantial evidence that a single release of commercially supplied convergent lady beetles reduced an aphid population on Elaeagnus. A lacewing release provided no evidence that C. rufilabris was effective in reducing aphids on stonecrop in landscapes. Also, a mixed species release of lacewings failed to produce a reduction in the abundance of the bean aphid on hawthorn.. Clearly, much remains to be learned before augmentative releases of lady beetles and lacewings can be used predictably and reliably by arborists, nurserymen, and landscapers to control aphids on landscape plants.
Arborists should use augmentative releases of natural enemies such as lady beetles and lacewings with caution. Based on these results, we cannot recommend release of lacewings for control of two species of aphids in landscape settings such as the ones described here: However, releases of the convergent lady beetle reduced aphid populations effectively in a landscape planting of elaeagnus and when beetles were confined to infested firethorn plants in containers. Several studies such as these must be conducted before augmentative releases can be used as a reliable management tool. Until such studies are conducted, arborists must rely on their own experiences with augmentative releases to decide if they are acceptable alternatives to other intervention tactics.
Acknowledgment
We thank The Smithsonain Institution for allowing us to evaluate predator releases in their landscapes. Financial support for this project was provided by the Competitive Grants Program of the Maryland Agricultural Experiment Station, International Society of Arboriculture Research Grant Program, and Horticultural Research Institute. Manya Stoetzel provided incomparable service in identifying the aphid species used in this study. This is Scientific Article Number A 6567, Contribution Number 8779 of the Maryland Agricultural Experiment Station.
- © 1994, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.