
Abstract
Mycoplasmalike organisms are obligate parasites of plants and insects. In recent years, advances have been made in detecting and identifying them, distinguishing among those associated with various plant diseases, assigning them to groups based upon homology of DNA base sequences, and determining their roles in decline of trees and shrubs. These advances are reviewed with special reference to ash yellows and elm yellows. Known and probable relationships among MLOs associated with various plant diseases are tabulated.
Mycoplasmalike organisms (MLOs) have been recognized as a distinct class of plant pathogens since 1967 (25), but their obligately parasitic habit has hindered research into their biology, ecology, and relationships to other organisms. Therefore, most diseases caused by MLOs remain poorly known, and those affecting trees and shrubs are generally uncontrolled. The rise of biotechnology has fostered advances in understanding of mycoplasmal plant pathogens. The purposes of this review are to highlight some of these advances and point out their significance for the eventual control of MLOs that affect trees and shrubs. Ash yellows and elm yellows will receive emphasis, because they are significant diseases of shade trees and subjects of the authors’ research.
Nature and Occurrence of Mycoplasmalike Organisms
MLOs are prokaryotes that lack cell walls and are among the smallest organisms known. They parasitize plants and certain insects that act as vectors. MLOs colonize plants systemically by way of phloem sieve tubes, which are the only cells commonly invaded. MLOs can be visualized in detail only by electron microscopy, but they can be detected in phloem by means of various microscopic, immunological, and recombinant DNA-based techniques. MLOs are considered to belong to the class Mollicutes and are named for their resemblance to culturable microorganisms in the genus Mycoplasma. Their genetic code is usually carried on a single chromosome that is smaller than the chromosomes of walled bacteria but comparable in length to those of culturable mollicutes (66). MLOs have not been formally classified and named at the genus and species levels, because the physiological and metabolic tests required for taxonomic descriptions of prokaryotes can only be done with pure cultures. Several contemporary reviews about plant and insect MLOs are available (17,47,58,76,93).
MLOs induce hundreds of plant diseases (76), of which over 150 affect woody plants. Disorders caused by MLOs are sometimes called “yellows diseases,” because the symptoms are similar to those of the much-studied aster yellows (50). These diseases are important primarily in trees and shrubs, because they are usually incurable and often kill or reduce the productivity of the plants (for reviews, see references 41,97, and 106). The most prominent of these diseases in North America include ash yellows (AshY) (71,105), bunch diseases of pecan and walnuts (70,97), elm yellows (EY) (54,100), lethal yellowing of palms (75,112), pear decline (49,77,115), and X-disease of Prunus species (48,88).
MLO Vector Relations
MLOs are vectored by over 100 species of insects, primarily leafhoppers, but also planthoppers, psyllids, and even a stink bug (24,83,111). Some vector species transmit MLOs associated with several diseases, and some MLOs have several vectors. For example, at least 16 leafhopper species transmit MLOs that cause X-disease of peach and cherries (88). Vectors ingest MLOs in phloem sap. The parasites multiply within various organs and cell types of a vector and eventually attain high concentrations in the salivary gland where they are in position to be injected into another plant. Adult vector insects are responsible for most transmissions into plants. MLOs do not infect vectors’ embryos. Therefore, infected plants have a crucial role in perpetuating the parasites.
Vector biology and habits have been elucidated for only a few of the most intensively studied mycoplasmal diseases of trees, e.g.,X-disease of Prunus species (12,31,88), pear decline (15,49,114), and lethal yellowing of palms (43). Little is known about the vectors, vector biology, or non-economic plant hosts of MLOs that affect other North American trees, or about epidemiology of the diseases.
Outlook
Fortunately, few MLO-induced diseases are characterized by devastating epidemics. Insidious weakening of infected plants is more common. In recentyears, MLOs have been found associated with several tree decline syndromes in the USA and in Europe, and more reports of these associations are likely. MLOs associated with slow decline syndromes probably interact additively with other stress-inducing factors, such as water shortage and defoliating insects. The main practical tasks for plant pathologists studying these diseases are to ascertain the role of MLOs among other stressinducing factors associated with tree decline and to identify plants that resist or tolerate MLO infection. The horticultural use of resistant or tolerant plants, either directly or as rootstocks, will minimize the future impact of MLOs.
Trees that tolerate MLO infection without soon undergoing irreversible decline may benefit from care designed to promote vigor or reduce stress. We advise owners of MLO-infected ash trees that do not show severe dieback to give the trees routine good care, because such trees are known to respond to year-to-year variation in water supply. Trees with severe dieback or affected by quick-decline syndromes such as that of elm yellows in American elm, on the other hand, are unlikely to respond to care. It is not essential to know whether or not MLOs are present in a given tree, because the diagnosis will not affect the prescription for care.
Destructive epidemics of MLO-induced diseases are likely to occur from time to time as MLOs in asymptomatic plants are introduced into new localities where susceptible plants and efficient vectors exist or as new vectors are introduced to existing MLO ranges. MLO introductions to the USA were apparently responsible for the outbreak of lethal yellowing of coconut palms in Florida (75) and could have caused the outbreak of EY in American elms.
Vector food preferences and habitat preferences determine which plant species in nature commonly become infected with MLOs. A change in food preference of an MLO vector could trigger an outbreak of a “new” disease caused by MLOs already present in other plant species. The broad experimental host range reported for western X-disease MLOs (44) illustrates that many plant species may be compatible hosts for a given MLO strain that does not commonly infect them.
Ash Yellows
Ash yellows occurs naturally in at least 12 indigenous and exotic species of Fraxinus in the USA and Canada but has not been reported from other parts of the world. Suscepts include Fraxinus americana (white ash); F. angustifolia (syn. F. oxycarpa); F. bungeana; F. excelsior (European ash); F. nigra(blackash); F. latifolia (= F. oregona, Oregon ash); F. ornus (flowering ash); F. pennsylvanica (green ash, red ash), F. profunda (= F. tomentosa, pumpkin ash); F. potamophila; F. quadrangulata (blue ash); and F. velutina (Arizona ash, Modesto ash, velvet ash) (39, 105).
AshY MLOs also induce lilac witches’-broom, a disease that causes decline in many cultivars of lilac and has devastated hybrid lilac collections in some arboreta (36,37,39). The relationship between ash and lilac MLOs was established by several methods: reciprocal graft transmissions which caused typical symptoms of the respective diseases; identical symptoms induced by ash and lilac MLOs in periwinkle to which the MLOs were transmitted by dodder; hybridization of DNA from MLO-infected ash and lilac with AshY-specific DNA probes; and restriction enzyme analysis of DNA fragments amplified by PCR from ash and lilac MLOs (32,39).
The principal reported range of AshY and lilac witches’-broom extends from Minnesota and Missouri to southwestern Quebec and Massachusetts (28,69,105). The disease also occurs in southwestern Utah in velvet ash (32,103). MLOs detected in Modesto ash in Arizona (8) and Nevada (105) have not yet been identified. Reports of ash witches’-brooms in southern USA have appeared (e.g., 86), but MLO infection of ash there has not been confirmed.
AshY is best known in white ash, in which it causes rootlet necrosis, slow growth, dieback, and sometimes death (26,71). Diseased trees of this species may display mild to severe chlorosis, undersized foliage, rosettes, and witches’-brooms (40,71). Diseased saplings sometimes die within 1-2 yr after growth reduction begins. Not all MLO-infected white ash sampled at a given time display suppressed growth, however (34,105). MLO-in-fected nondeclining trees may be tolerant of MLO infection or may be in early stages of susceptible reactions.
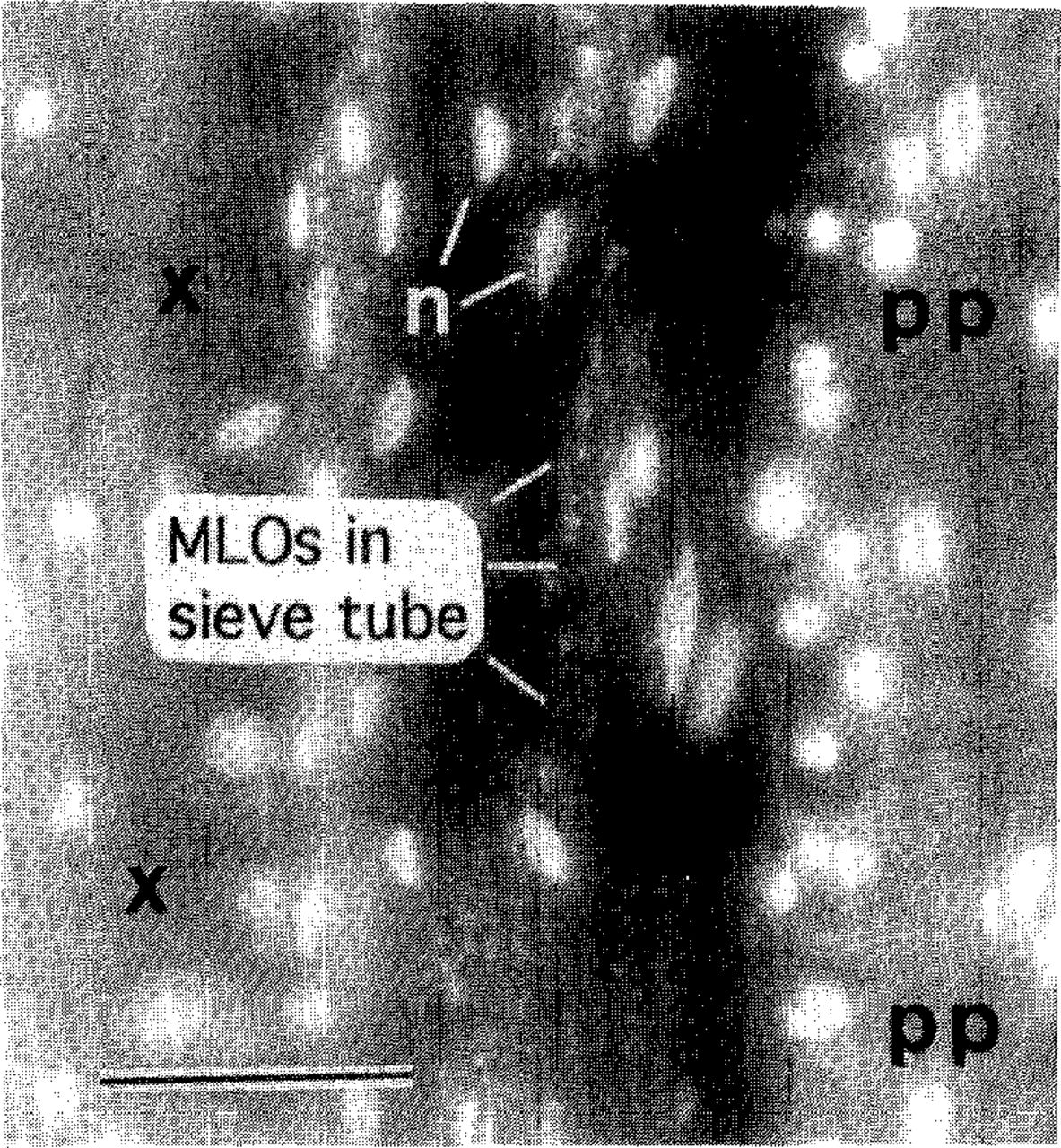
Field diagnosis of AshY is based upon detection of witches’-brooms. Other symptoms of the disease are often uninterpretable, because causal factors other than MLOs can induce similar symptoms. Unfortunately, only a small proportion of ash affected by AshY have witches’-brooms at a given time. Therefore, unambiguous diagnosis usually requires detection of MLOs by a microscopic test such as the DAPI fluorescence test (Fig. 1). The results may be corroborated, if desired, by tests that specifically identify AshY MLOs: DNA-DNA hybridizations, MLO DNA amplification by polymerase chain reaction (PCR) followed by restriction enzyme analysis of the PCR product, or immunological tests using an AshY-specific antibody (20,32,39). These tests have revealed that MLO strains from ash and lilac across the range of AshY are all closely related to each other and differ from strains detected in other plants (32,39).
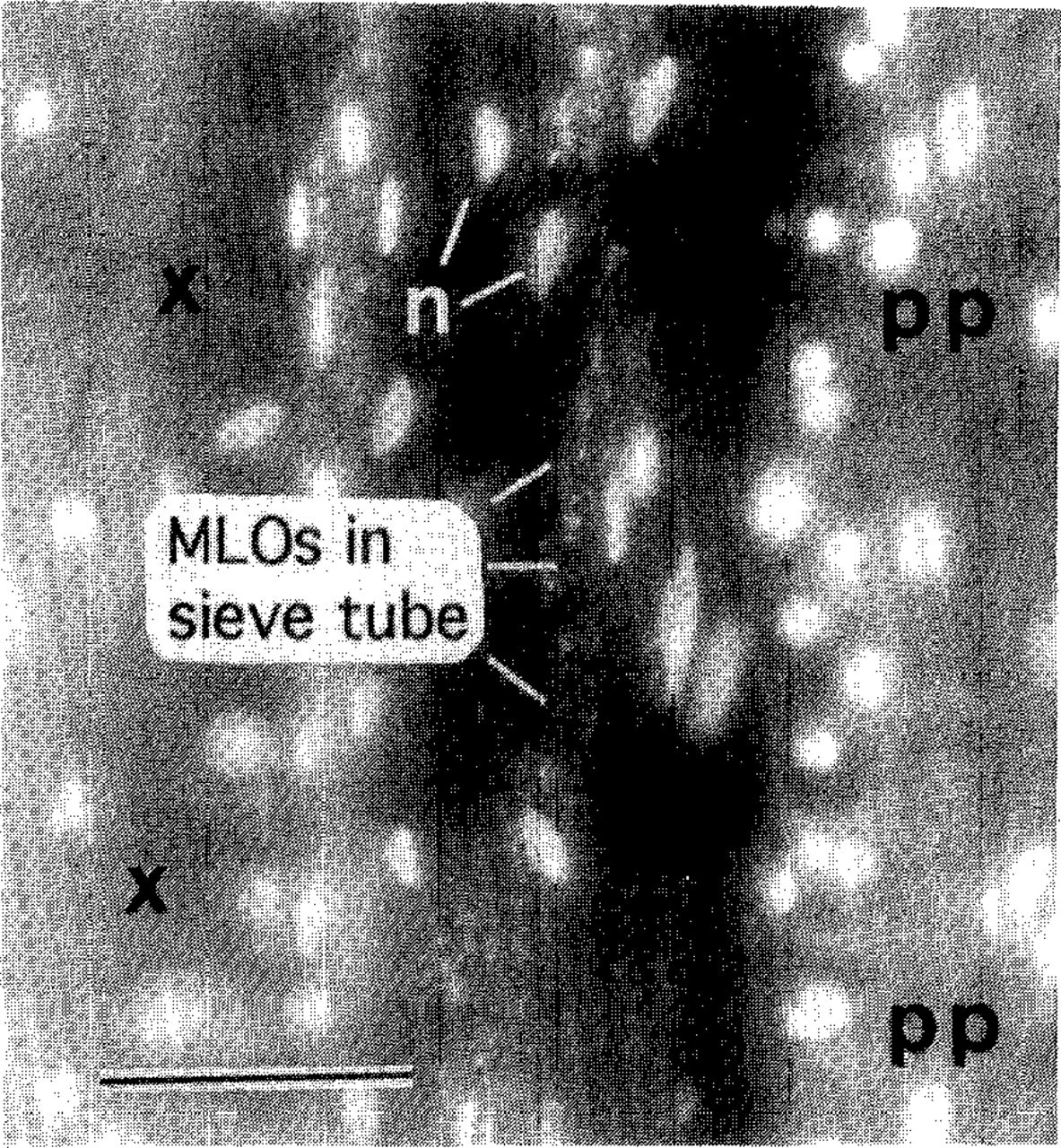
DAPI test for MLO detection. A longitudinal section of white ash rootlet was treated with DAPI (4’,6-diamidino-2-phenylindole·2HCI) and observed by means of fluorescence microscopy. Brightly fluorescing objects (n) are plant nuclei. Xylem (x) and phloem fibers (pp) are autofluorescing. DNA of MLOs in phloem sieve tubes appears as fluorescent speckles. Sieve tubes, which lack nuclei when mature, are otherwise invisible. Bar = 50 μm.
AshY in eastern North America is most common and severe where wooded lands occur in patches (105). The disease occurs infrequently in areas covered primarily by forest (105,109). AshY in white ash is believed responsible for much of the damage to which the name “ash dieback” (38) was applied. However, ash trees unaffected by MLOs often display progressive loss of vigor and dieback, presumably induced by adverse environment or insect attack (69,103,105).
In green ash, AshY causes growth suppression (102), abnormally upright branching habit in young trees, and formation of witches’-brooms. MLO-associated dieback in this species seems restricted to saplings and small trees with crowns beneath the canopy of other trees. A survey in four midwestern states revealed dieback of this species in forest stands to be common but not consistently associated with MLO infection (69). The dieback was thought due to episodes of drought during the 1980s. Green ash infected with AshY MLOs does not sustain rootlet necrosis to the extent that white ash does (26), which may explain the lack of prominent MLO-associated dieback.
In velvet ash studied in southwestern Utah, AshY MLOs usually cause no symptoms unless infected trees are near death from other causes such as water shortage or defoliation by insects (103). In severely debilitated velvet ash, the MLOs may induce witches’-brooms and dense clusters of dwarfed, simple-leaved shoots low on the trunk and at the root collar. Tolerance of MLO infection by velvet ash was indicated by lack of MLO-associated growth reduction in infected wild trees and by vigorous growth of infected trees in greenhouse tests. White ash inoculated with the same strain of MLOs developed rootlet necrosis and stunted shoots (103).
Experience with AshY in other ash species is too limited to permit comments about their relative susceptibilities to MLO-induced damage or possible tolerance of infection. Nothing is known about possible resistance of ash species and cultivars to colonization by AshY MLOs. Epidemiology of AshY has not been studied in detail. Incidence of “ash decline” increased overtime on plots in New York State (9), but MLO association with the decl ine was not studied. We have recorded annual increases of 0-10% in AshY incidence in young populations of white ash in central New York State (Sinclair and Griffiths, unpublished). These rates of disease increase, if sustained for 10 years or more, would be sufficient to result in high incidence and major impact on ash volume.
AshY MLOs are probably transmitted only among and between ash and lilac plants in nature, because these MLOs were not detected in other plants during a search for alternative hosts on sites where AshY was prevalent in ash (32). The identities of AshY vectors are still unknown. A preliminary report of vector ability of a leafhopper and a spittlebug (72) has not been corroborated.
Elm Yellows
Elm yellows kills highly susceptible elms within a year and causes witches’-brooms and decline in more tolerant elm species (100,106). Once known only in midwestern USA, EY has spread into eastern states and southern Ontario (72,73). Destructive epidemics have occurred south of latitude 43° N, approximately (54,72,100), and the disease is considered a threat to native elms in eastern North America south of that latitude. Although spot occurrences of EY have been recorded farther north (e.g., 110), epidemics are not sustained there, perhaps because vectors do not persist.
EY also occurs widely in Europe, where reports of witches’-brooms and MLO-associated decline of elms date from 1981 (14,85). The diagnosis of EY in Europe was first made on the basis of similarity of the symptoms to those described for graft-inoculated Eurasian elms in New York State (7). Recently, DNA hybridization tests revealed that elm-inhabiting MLOs in Germany, Italy and the USA are all members of a discrete group (61,74). Natural infection of Chinese elm by EY MLOs in New York State was also verified (61,74). The diseased trees developed witches’-brooms and dieback.
New elms resistant to Dutch elm disease (DED) are being deployed in the USA. These elms represent both Ulmus amerícana and hybrids and selections of Eurasian origin. The DED-resistant American elms, such as those of the ‘American Liberty’ group (108) are fully susceptible to EY (Sinclair, unpublished). One municipality in New York State planted a number of DED-resistant American elms as street trees, only to lose them to EY. The reactions of Eurasian hybrid cultivars to EY MLOs have not been well characterized, but these cultivars are thought to be resistant or immune because Eurasian species growing in the vicinity of American elms in the USA escaped epidemics of EY that killed the latter species during the 1930s through 1970s. Recent observations of EY-induced decline in Eurasian elm species in Italy, however, indicate that resistance or tolerance of new DED-resistant elm cultivars to EY should be evaluated critically (78).
EY epidemics in highly susceptible American elms progress rapidly until all large individuals of this species in the affected localities are dead (54,100). Saplings often escape infection until their crowns grow above low vegetation, but then they too are killed. In central New York State, where an EY epidemic began in 1971, the disease is prominent every summer in saplings that have grown since the large trees died. EY apparently spreads only from elm to elm, because we have searched unsuccessfully for EY MLOs in other plants growing on sites of EY epidemics (32).
One vector of EY, the leafhopper Scaphoideus luteolus, is known from research in Ohio (2), but the vector(s) responsible for outbreaks of EY in northeastern USA and in Europe are unknown. A report of EY transmission by the leafhopper Allygus atomarius and the meadow spittlebug, Philaenνs spumarius (72) remains unconfirmed.
Recent Advances
Phylogeny and classification of MLOs
Despite inability to obtain pure cultures, researchers using techniques of molecular biology have made progress differentiating MLOs, determining their relationships to one another and to other kinds of microorganisms, and constructing a preliminary classification (33,51,58,63,67,81,82,91). MLO genes that encode ribosomal RNA and ribosomal protein have been detected through comparisons of their base sequences with those of the corresponding genes of culturable prokaryotes (51,64-67,82). These comparisons have revealed that the closest known relatives of MLOs are mollicutes in the genus Acholeplasma (51,67,82).
MLOs are genetically diverse. This diversity has been studied using combinations of three general methods: a) DNA-DNA hybridizations using probes derived from MLO DNA (e.g. 53,59,61,74), b) analyses of base sequences of small fragments of MLO DNA cloned in bacteria or amplified from diseased plant or insect DNA by polymerase chain reaction (18,91), c) analyses of MLO DNA fragments obtained by digestion with restriction endonucleases (53,59,62,63,91).
MLOs are now grouped on the basis of similarities and differences in their DNA sequences (58,63,81,91) (Table 1). Two methods have been used to determine these groups. One involves delineation of MLO “strain clusters” based on DNA-DNA hybridization analyses using randomly cloned DNA probes derived from various MLOs (17,20,58,59,61,62,79). Each cluster contains MLO strains that share extensive DNA sequence homology with one another and are distinct from strains in other MLO strain clusters. Strains within a given cluster are differentiated by restriction enzyme analysis of each MLO’s chromosomal DNA. In this analysis, MLO DNA is digested with enzymes that cut the molecules at sites where specific base sequences occur. Different MLOs vary in recognition sites for a given restriction enzyme and thus in the numbers and sizes of DNA fragments resulting from digestion. The DNA fragments are separated according to size by gel electrophoresis and later are tested for ability to hybridize with selected DNA probes (16,53,59,62). Each probe hybridizes only with DNA fragments that have base sequences homologous to those of the probe. MLOs are differentiated or identified by comparing the hybridization patterns (4,5,16,59,62,74,99).
Differentiation and classification of mycoplasmalike organisms on the basis of DNA sequence analysisa
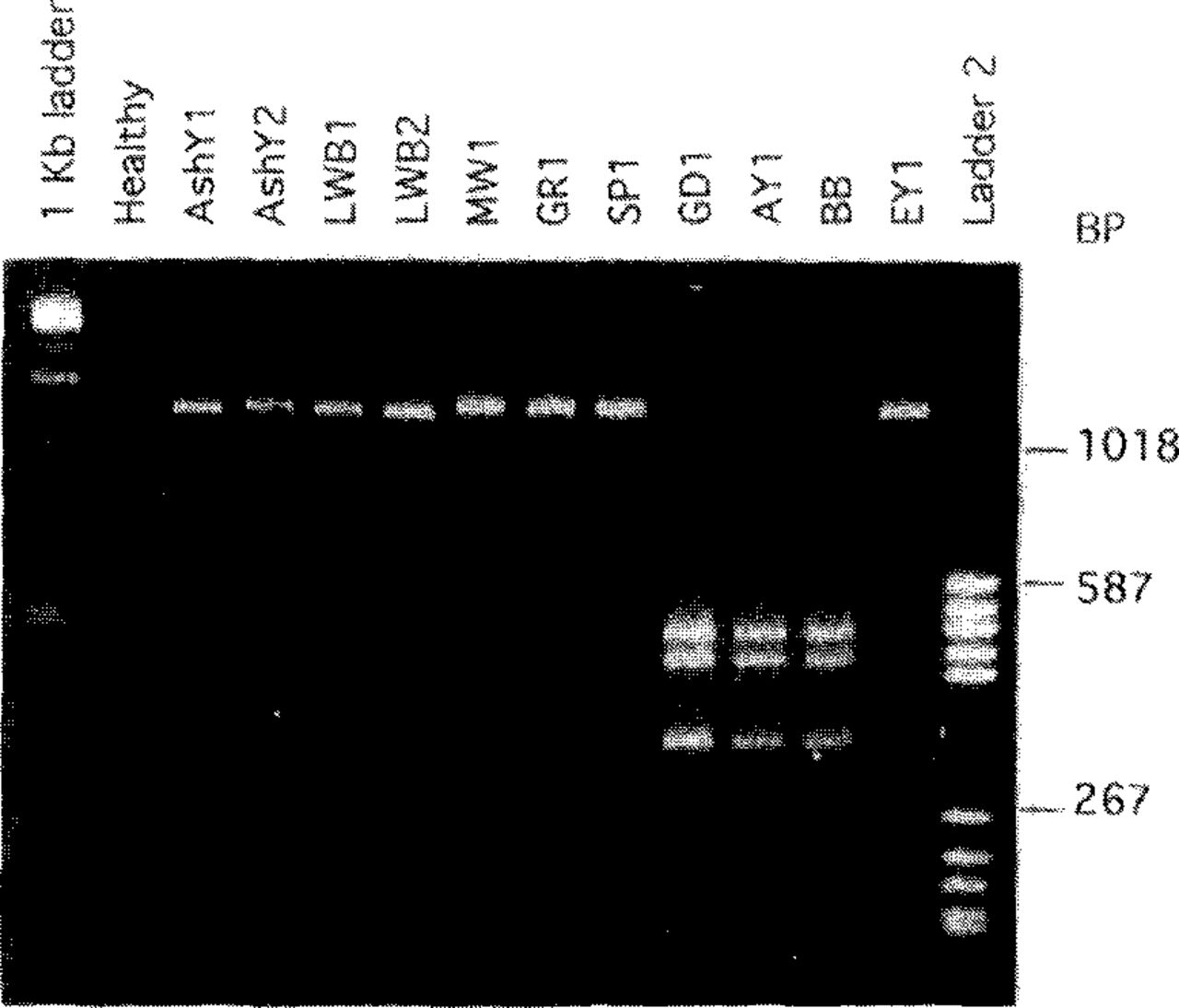
The second approach to MLO classification involves comparative analyses of nucleotide sequences in the l6S ribosomal RNA gene (1,18,19,23,27,63,81,87,91). The expression 16S refers to a particular subunit of the ribosome. These sequences are highly conserved, i.e., relatively stable over long time spans. Therefore, differences in these sequences between MLO strains are likely to reflect evolutionary divergence. For analysis, DNA is extracted from MLO-infected plants or insects, and a portion of the 16S ribosomal gene that is common to all MLOs but not possessed by other organisms is amplified (replicated many fold) by PCR (1,22,63). The sequence of nucleotides in the amplified DNA is then examined, usually by restriction enzyme analysis. In this procedure, samples of the DNA are digested with different restriction enzymes, and the fragments obtained with each enzyme are separated by gel electrophoresis. The numbers and sizes of the fragments vary among MLO groups and strains. To differentiate and identify MLO strains, data obtained with each of several enzymes for a given strain are recorded and compared with data for other strains. Figure 2 shows an example involving just one restriction enzyme.
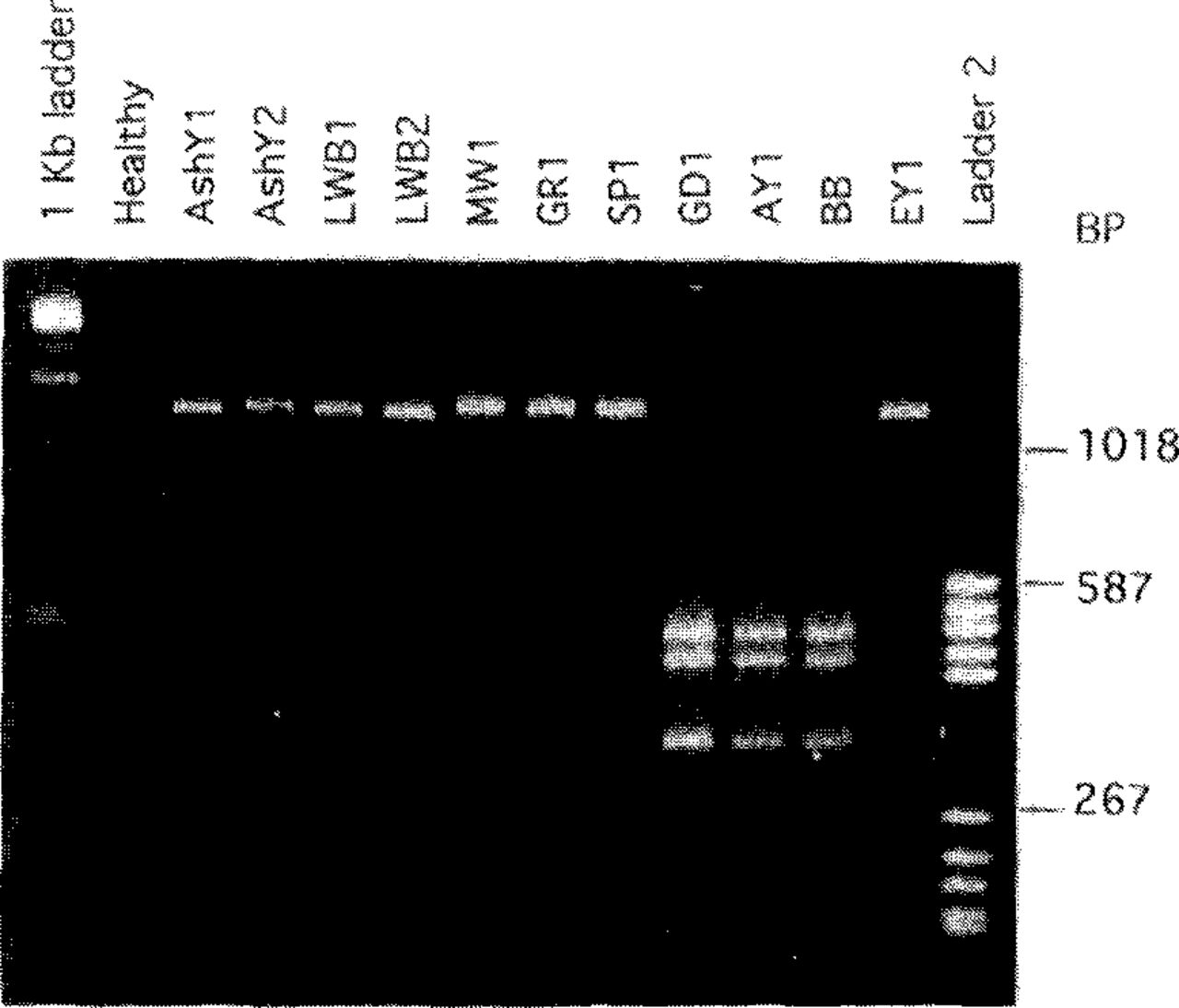
MLO differentiation by restriction enzyme analysis. A segment, approximately 1200 base pairs (BP) long, of the 16S ribosomal RNA gene of each of several MLO strains was amplified by polymerase chain reaction (PCR). PCR products were digested with restriction enzyme Kpnl and subjected to electrophoresis in an agarose gel, then were stained with ethidium bromide and photographed under UV illumination. The amplified DNA segment from eight of the MLO strains was not cut by this enzyme, so a single band of DNA from each of these strains is visible in the gel. MLO strains belonging to the aster yellows cluster (GD1, AY1, and BB) have two recognition sites for KprΛ in the amplified DNA segment, resulting in three fragments after digestion and distinguishing these MLOs from the others tested. MLO designations: AshY-ash yellows, AY-aster yellows, BB-tomato bigbud, EY-elm yellows, GD-dogwood stunt, GR-goldenrod yellows, LWB-lilac witches’-broom, MW-milkweed yellows. “Healthy” refers to DNA from healthy periwinkle (no PCR product obtained). Outside lanes are molecular size standards.
Three of the preliminary classification schemes shown in Table 1 are based on 16S ribosomal gene sequence data. These classifications are in general agreement with that based on DNA-DNA hybridization analyses. More MLO groups and strain clusters will probably be designated as additional strains are studied.
Genetic versus phenotypic variability in MLOs
Genetic differences among MLO groups and strains, although easy to detect, can not yet be related to specific MLO characteristics or behavior. Research in this area is limited because MLOs can not be grown in pure culture. Therefore, it is not yet possible to manipulate MLO genes in ways that would permit identification of those coding for characters such as virulence or plant or insect host range.
Extrachromosomal DNA, presumably of plasmids (circular DNA molecules that replicate independent of chromosomal DNA), has been detected in various MLOs and seems to be characteristic of some strains, notably members of the aster yellows cluster (52). The function(s) of plasmid DNA in MLOs are unknown, but the loss of plasmid DNA by a clover phyllody MLO during micropropagation of its plant host was associated with loss of ability of the MLO to be transmitted by its leafhopper vector (21). The implication of this finding is that important biological properties such as vector specificity may be gained or lost with plasmid DNA.
Phenotypic differences among MLOs can, at present, be detected only in terms of MLO interactions with plants or insects. For example, MLOs from various plant species can be transferred, via dodder (Cuscuta spp.), into a standard plant host, such as periwinkle, in which symptoms induced by various strains can be compared.
MLO groups vary in degree of host specialization. For example, MLOs in the ash yellows cluster apparently affect only species of ash and lilac in nature (32,39), while members of the aster yellows cluster infect a broad array of plants. Plant host ranges are not presently useful for distinguishing MLO strains, because host range data are available for few strains. Moreover, the broad experimental host ranges reported for some MLOs (e.g., aster yellows and X-disease MLOs) reflect the fact that certain vector species will accept diverse plant species as food sources if preferred species are unavailable. MLOs can probably multiply in phloem sap of many plants to which they would not be introduced by vectors in nature.
Vector specificity varies among MLO strains, but its usefulness for distinguishing them is limited by lack of knowledge of the vectors of many diseases and difficulty in acquiring and handling insect colonies. Moreover, the detection of MLOs within an insect does not necessarily indicate it is an MLO vector, because insects can become infected with MLOs that they do not transmit (89,107).
Methods for MLO detection and identification
Field diagnosis of MLO diseases depends upon recognition of specific symptoms such as witches’-brooms. Unfortunately, MLO-infected plants may fail to display diagnostic symptoms. Therefore, microscopic examination and/or molecular tests are often necessary to supplement field observations or corroborate a diagnosis. These tests are currently performed only in research laboratories.
The DAPI fluorescence test (92) (Figure 1) is widely used for nonspecific detection of MLOs in plants. Nonspecific detection by DNA probes or PCR has also been achieved in several laboratories (1,3,22,27,63,74,81). The latter procedure is preferred because of its sensitivity. MLOs can be identified by means of DNA hybridizations (4,5,16,20,32,35,39,61,62,74,79,99,113), analyses of PCR products (3,19,32,57,59,61,81,87), or immunological tests (6,13,29,32,45,60,68). Most recently, PCR primers are being designed that amplify 16S ribosomal DNA of only those MLOs belonging to particular MLO clusters, such as the aster yellows cluster (18). Application of these molecular techniques to MLO identification in naturally infected plants and insects is just beginning.
Changing Concepts
For many years, concepts about MLO-induced plant diseases were derived mostly from the realm of plant virology, because viruses were blamed for diseases now known to be caused by MLOs. New knowledge about MLOs and diseases that they cause has reinforced some concepts and caused others to be modified or discarded.
The concept that each disease is caused by one primary causal agent (i.e., a discrete group of closely related MLO strains) is not generally applicable. The MLOs associated with certain diseases, ash yellows and elm yellows for example, seem to represent discrete groups, but different MLOs may cause nearly indistinguishable symptoms in some plants. For example, decline of peach trees in North America is caused by MLOs of the X-disease cluster, but MLOs belonging to the aster yellows cluster and to a group of fruit tree-infecting M LOs that differ from X-M LOs are associated with peach decline in Europe (91). MLOs belonging to three distinct groups have been detected in grapevines with yellows symptoms (87) (Table 1).
The concept of inevitable decline of trees and shrubs infected by MLOs is correct but misleading, because decline is a phase of the normal life cycle. MLO-infected trees that are tolerant of infection may continue to grow satisfactorily, responding to environmental stimuli in the manner of healthy trees. We obtained evidence of such responses during studies of radial growth of green ash and white ash in New York State and velvet ash in Utah as affected by ash yellows. In all three species, year-to-year growth trends of MLO-infected trees were similar to the trends of healthy trees (102). However, the amount of annual growth of MLO-infected trees of the two eastern species was less than that of healthy trees, reflecting their lesser tolerance of infection. Highly susceptible white ash trees and occasional individuals of the less susceptible green ash affected by ash yellows undergo progressive growth decline accompanied by branch dieback, but many MLO-infected trees of these species maintain moderate growth and acceptable appearance for many years.
The concept that MLOs may have symptomless hosts remains valid and important, not only for understanding epidemics of MLO-induced diseases but for the potential of disease control. MLOs have been detected in asymptomatic as well as declining alders in Europe (56), in both declining and apparently healthy velvet ash in Utah (103), and in symptomless apricots (48) and ‘Peerless’ almonds (113) in California. The alder MLOs in Europe were found to be closely related to elm witches’-broom MLOs (74). Whether MLO strains are naturally transmitted between elm and alder is unknown, however. The MLOs in velvet ash in Utah were identified as members of the ash yellows cluster (32), and grafting experiments revealed that velvet ash is tolerant of infection by these MLOs (103). The almond MLOs in California, which were found to belong to the X-MLO group, were detected only after bud grafts of almond on plum rootstock failed. The MLOs from tolerant almond induced necrosis at graft unions. Observations such as these reveal that MLO-infected symptomless plants can serve as reservoirs of MLOs that cause damage to susceptible plants. On the other hand, MLO-tolerant plants may be useful as rootstocks, as has been demonstrated for suppression of pear decline symptoms (95,115).
The concept of discontinuous MLO populations within diseased trees remains valid and explains variable symptoms and diagnostic results. For example, Seemüller et al. (95) detected MLOs in only 13% of diseased ‘Hardy’ pears on a quince rootstock, versus 81% detection in the same pear cultivar on a pear rootstock. Discontinuous MLO distribution was confirmed by results of graft transmission trials. The frequent confinement of pecan bunch MLOs to witches’-brooms has been demonstrated by pruning and grafting experiments and electron microscopy (summarized in reference 96). The MLOs associated with walnut witches’-broom could not be detected in all parts of a tree by means of DNA probes (11). Willow witches’-brooms are localized on affected trees, and healthy plants have been propagated from healthy-appearing parts of such trees (42, Sinclair, unpublished). Ash yellows MLOs were detected by means of the DAPI procedure more frequently in roots than in twigs of diseased white ash (10,101,104). MLO population levels also vary seasonally in diseased trees. Ash yellows MLOs and walnut bunch MLOs were detected more frequently in summer than in other seasons (10,11,101).
The concept that MLO infections are permanent and incurable should be accepted as generally true, but some exceptions occur. MLO populations associated with apple proliferation apparently die out in certain resistant Walus cultivars (46). Spontaneous remission of pear decline symptoms was observed in pear trees in Oregon and Colorado, apparently because the MLOs died during winter (114). Similarly, we have observed occasional spontaneous remission of ash yellows symptoms in young potted plants of white ash. Such plants, initially stunted and harboring MLOs that were detected by means of the DAPI test, began to grow normally after a dormant period at 0-2°C.
Scientific advances in plant mycoplasmology will probably be incremental until the development of a reliable method for growing MLOs in pure culture. This breakthrough seemed imminent in the early 1970s, but after numerous failures, most MLO investigators turned to other lines of work. Pure culture of MLOs will permit molecular-level studies of their interactions with plants and insects; delineation, naming, and classification of species; and genetic engineering of plants and MLOs that may lead to effective long-term controls.
Acknowledgments
This paper is based on a presentation to the International Society of Arboriculture at its 69th annual conference, Bismarck, ND, 9 August 1993. The authors’ research reviewed here was supported by the Cooperative State Research Service of the U.S. Department of Agriculture through the McIntire-Stennis Cooperative Forestry Research program and the Competitive Research Grants Office, grants 89-37151-4557 and 91-37303-6427, and by the U.S. National Park Service through contract CA-1463-5-0001. We thank George W. Hudler and Craig R. Hibben for helpful reviews of the manuscript.
- © 1994, International Society of Arboriculture. All rights reserved.
Literature Cited
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.
- 81.↵
- 82.↵
- 83.↵
- 84.
- 85.↵
- 86.↵
- 87.↵
- 88.↵
- 89.↵
- 90.
- 91.↵
- 92.↵
- 93.
- 94.
- 95.↵
- 96.↵
- 97.↵
- 98.
- 99.↵
- 100.↵
- 101.↵
- 102.↵
- 103.↵
- 104.↵
- 105.↵
- 106.↵
- 107.↵
- 108.↵
- 109.↵
- 110.↵
- 111.↵
- 112.↵
- 113.↵
- 114.↵
- 115.↵
In this issue








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.