
Abstract
Water use and water relations of sweetgum growing in an urban canyon were investigated. Predawn water potential, dawn-to-dusk stomatal conductance, and leaf morphology were measured over a two-year period at a downtown site in Seattle, Washington. Trees received four hours of direct midsummer sunlight during midday. This was compared to similar-aged sweetgum street trees growing in a neighboring park-like setting. Specific leaf area and leaf presentation angle of the canyon trees were characteristic of shade acclimation. Stomata of the canyon trees quickly opened in response to light during the morning shade period, but closed rapidly in response to low humidity during the sunlit period, and remained closed through the afternoon shaded period. Overall lower conductances and leaf temperatures in the canyon trees resulted in transpiration rates that were lower than the park trees. The data suggest that trees growing in urban canyons will deplete soil water less rapidly due to the effects of reduced irradiance on tree transpiration.
Knowledge of microclimates is an important part of the ongoing effort to characterize the interaction between the urban environment and trees. Urban canyons are microclimates that have limited gap-like inputs of direct-beam solar radiation due to horizon topographies that periodically obscure the sun (8,9). We previously reported that the dominant effect of urban canyon microclimates on trees appears to be related to low irradiance (4). Leaf morphology of sweetgum street grees growing in an urban-canyon were consistent with shade acclimation typically observed in understory trees (4, 5). Since radiation also drives evapotranspiration, low irradiance in urban canyons has implications for tree water use and water relations. Forest trees acclimated to low-irradiance understory conditions exhibit less favorable water relations when exposed to high irradiance in an overstory gap (12). Given the dynamic nature of forest gaps, such a response will be short lived as the overstory gap fills in. In contrast, the static gap of urban canyons imposes a mosaic of periods with heavy shade and sudden, high-intensity irradiance gaps where air temperature and vapor pressure deficit increase rapidly (8). How shade-acclimated foliage makes the transition between chronic high and low irradiance in terms of water use and water relations is uncertain. This paper reports on water use and water relations of sweetgum growing in an urban canyon and park.
Methods
Sites
Two sites, a full-sun park and an urban canyon, previously described in detail (8), were investigated. The full-sun park site was a city block, on a major east-west street in a commercial-residential district in Seattle, Washington. It was bordered on the north by a municipal park with extensive, well-maintained turf. The canyon site was a city block situated on the east side of a major north-south street in the central business district. Adjacent building heights ranged from 50 to 100 meters (150-300 ft). Ten-year-old sweetgum (Liquidambar styraciflua) street trees (n=10-12) of similar size, located in the parking strip between the curb and sidewalk, were studied at each site in 1986 and 1987. The canyon trees were in a 1.3 x 1.3 m opening in the cement parking strip contiguous with the sidewalk, while the park trees were in a turfgrass parking strip. The park trees received supplemental water whenever municipal watering of the turf occurred. Three trees in sequence at each site, at the north end of the canyon block and the east end of the park block, were selected for intensive study.
Data collection
Dawn-to-dusk stomatal conductance, wet bulb and air temperatures, and photosynthetic photon flux density (PPFD) shortwave radiation were spot-measured sequentially at each site on three dates each in 1986 and 1987. Five representative full-sun leaves at mid-crown were selected forconductance measurement with a steady-state porometer (Li-Cor Inc., Lincoln NE); in 1987 leaf temperature was measured on the same leaves with a fine-wire, ventilated thermocouple. Radiation and PPFD were measured with a pyranometer and quantum sensor (Li-Cor Inc. Lincoln, NE), respectively, leveled at 2 m. Temperatures were measured with a shielded, fan-aspirated psychrometer adjacent to the trees at each site, from which vapor pressure deficits (VPD) were calculated. Predawn water potential was determ i ned over both seasons with a pressu re chamber (Soil Moisture Inc. Santa Barbara, CA) on the terminal 10 cm of a single randomly-selected shoot collected at mid-crown level from each tree. Reference evaporation was measured over the 1987 season from a USDA Class-A evaporation pan located on the University of Washington campus approximately 5 km northeast of the study sites.
Two shoots from primary scaffold branches at mid-crown of each tree, one each from the north and south sides, were harvested in fall 1987. Prior to leaf removal, angle of presentation (the angular deviation of the mid-rib of the leaf blade from horizontal, 90°=vertical downward), was measured (with the excised branch repositioned at the in-situ angle. Specific leaf area was calculated as the leaf area (determined with a leaf area meter) divided by the weight of each leaf after drying at 60°C for 24 hours. Specific leaf area and leaf angle were determined for every leaf on each of the two shoots. Crown silhouette (projection) areas were determined in midsummer 1987 as the product of shaded ground area (measured from the number of shaded intersections on a 0.2 m-spaced grid) under each tree at midday and the sine of solar elevation.
Data analysis
Dawn-to-dusk stomatal conductance from both seasons was regressed on VPD measured concurrently; measurements below 200 W/m2 irradiance were eliminated because of confounding light effects (10). For the canyon trees this limited the data set to measurements taken during the sunlit period. In addition, dawn-to-dusk conductance from the morning and afternoon shaded periods at the canyon was regressed on the corresponding PPFD level. Leafair temperature difference was plotted against air temperature to evaluate possible trends. Transpiration was calculated for trees at both sites from data collected during dawn-to-dusk studies on August 4, and August 18,1987. Instantaneous transpiration (g/m2/s) for each cycle through the day was calculated as the product of stomatal conductance (m/s) and vapor density difference (g/m3) between the leaf at saturation (at leaf temperature) and ambient vapor density. The product of instantaneous transpiration and time in seconds in each sampling period summed through the day gave total daily transpiration, and was then converted to mm.
Results and Discussion
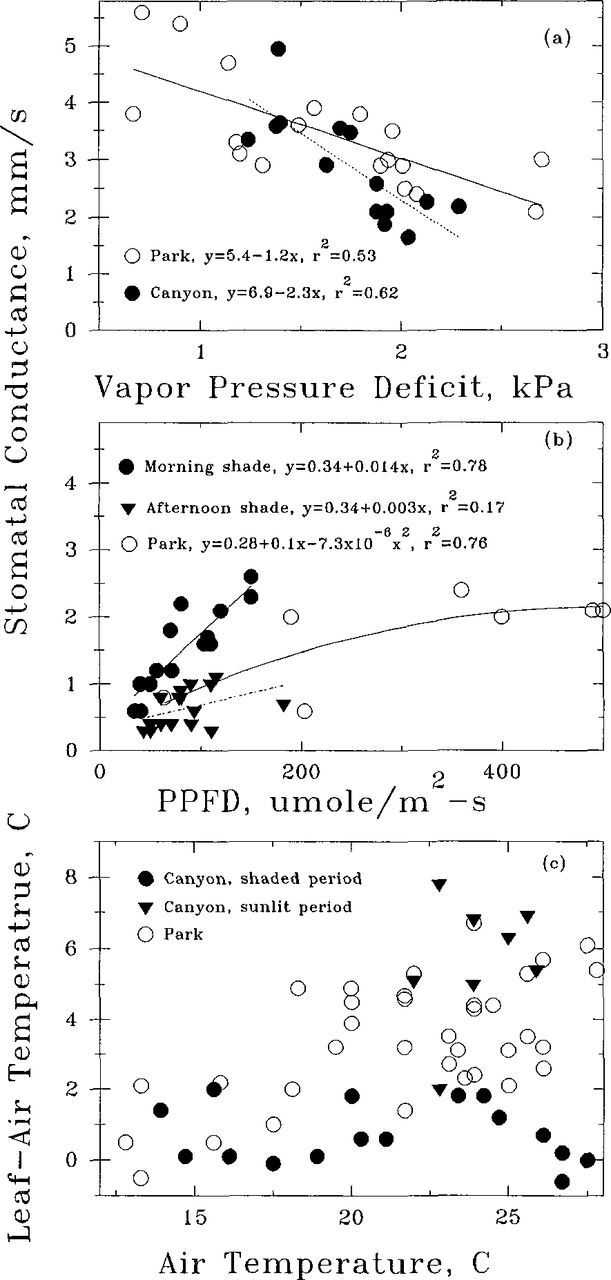
The canyon trees had higher specific leaf area (more leaf area per gram of leaf) and a more planar angle of leaf presentation, 58 versus 83 degrees, than the park trees (Table 1). Greater specific leaf area and more planar leaf orientation of the canyon foliage are characteristic of shade acclimation normally found in trees grown in the forest understory (3,7). Shade acclimated foliage resulted in stomatal behavior and a leaf temperature different from that of the park trees (Fig. 1). Conductance of both canyon and park trees were sensitive to vapor pressure deficit, declining as VPD increased (Fig. 1a). Stomata of the canyon trees appeared to be more sensitive, as the slope of the response was nearly twice as great as that of the park trees. Stomatal sensitivity is a common response in many woody plants that moderates water loss (6), and it would reduce transpiration in the canyon trees during the sunlit gap periods.
Specific leaf area, leaf angle of presentation, crown silhouette area, and transpiration of sweetgums growing in park and canyon sites in Seattle, Washington. Values represent the average of three trees(± standard error).

Relationship between a) stomatal conductance and vapor pressure deficit of park and canyon trees during direct sunlight; b) stomatal conductance of canyon sweetgum under diffuse-sky photosynthetic photon flux density (PPFD) during morning and afternoon shaded periods, and park conductance collected when PPFD levels were below 400. Slopes of fitted curves were significantly different from zero at P=0.05; c) leaf-air temperature difference for park and canyon foliage with increasing air temperature. Data points in each graph represent the mean of three trees.
Stomatal conductance of the canyon foliage trees was sensitive to solar radiation during shaded periods (Fig. 2). The response varied between morning and afternoon. During morning shade, stomata of the canyon foliage opened rapidly in response to small increases in sunlight, while park conductance was less responsive at similar light levels which was more typical of sun foliage. Conductance of the canyon trees reached 50% of the daily high rate at irradiances emanating from diffuse-sky and reflected radiation that was 10% of full sun. Similar responses have been observed in stomatal sensitivity to ephemeral sunflecks by trees growing in the understory (13). During the afternoon shaded period, stomata of the canyon foliage were insensitive to changes in light levels. Canyon conductance generally remained below 1 mm/s at light levels less than 10% of full sun, in contrast to the higher morning rates at similar light intensities This persistent afternoon closure suggested some internal factor changed during the gap period and prevented re-opening.
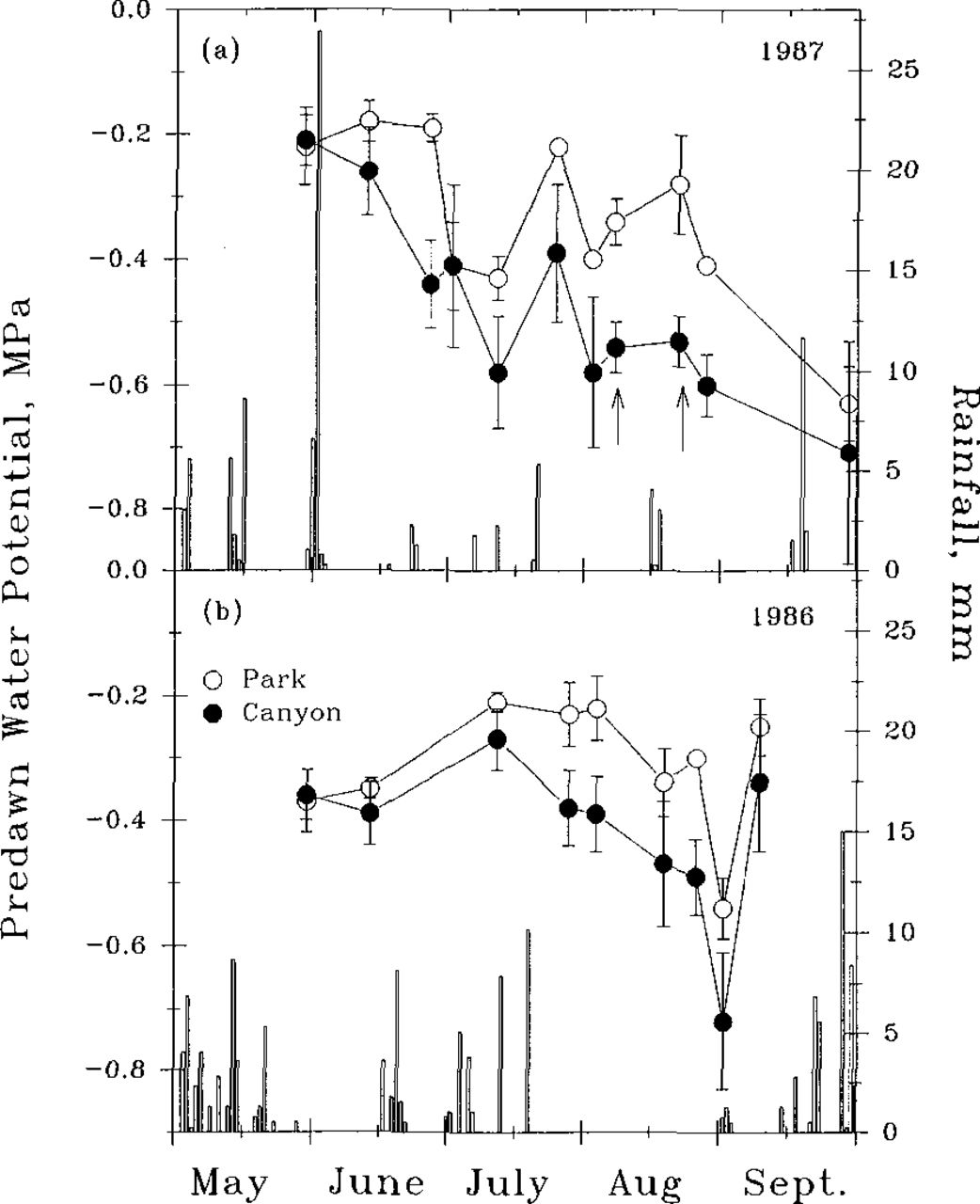
Seasonal changes in predawn leaf water potential for canyon and park trees during 1986 and 1987. Data points represent the average of three tree observations per site and the standard error. Data points without error bars have error terms too small to be shown. Arrows indicate the dawn-to-dusk study dates used in calculating transpiration rates in 1987.
Leaf-airtemperature differences varied between the park and canyon trees (Fig. 1c). The park trees exhibited an increasing difference as air temperature increased towards 30° C. This was due to stomatal closure at the correspondingly higher vapor pressure deficits associated with higher air temperatures. Leaf-air temperature difference of the canyon foliage was strongly affected by irradiance. During the sunlit period foliage temperature of the canyon trees was 5-7 degrees warmer than that of the air. This was slightly greater than that of the park trees at similar air temperatures and could have been due either to greater stomatal closure in response to increased VPD, or possibly to more horizontal leaf orientation intercepting more sunlight. In contrast, during the shaded periods the lack of radiation dropped foliage temperature of the canyon trees substantially. Canyon foliage temperature difference was only 1-2°C warmer than the air temperature even during afternoon shade periods when air temperatures reached 30°C.
Lower overall stomatal conductances and leaf temperatures resulted in lower calculated transpiration rates in the canyon foliage than in the park trees (Table 1). On August 4 average daily transpiration of the park trees was nearly two and a half times as great as the canyon trees. On August 18 transpiration declined at both sites due to lower conductances and a shorter day, but the decline was greater in the park trees. The park trees still had slightly highercalculated transpiration rates than the canyon trees. Transpiration rates were not a constant fraction of reference pan evaporation, which was nearly the same on both dates, 3.9 mm on August 4, and 3.8 mm on August 18.
Overall silhouette projection area of the canyon trees was greater than the park trees (Table 1). Taking the product of the calculated transpiration rates, based on m2 of leaf area, and crown size gives an estimate of total tree transpiration in volumetric units. On a whole-tree volumetric basis, transpiration would have still been greater on August 4 for the park trees, but similar between the two sites on August 18.
The canyon trees exhibited only slightly greater water stress than the park trees despite not receiving any irrigation (Fig. 2). Seasonal changes in predawn water potential over both seasons showed the park trees to have less negative levels than the canyon trees, a result of supplemental watering. The differences were more pronounced during dry periods from mid-July to late August during both years. Maximum differences between trees at the two sites that never exceeded −0.3 MPa over both years indicated that the canyon trees were not substantially stressed for water. Tolley and Strain (11) found delayed onset of water stress when sweetgum seedlings were grown under low irradiance as compared to high irradiance. The reduced input of solar radiation for the canyon trees apparently reduced transpiration rates sufficiently low so that more leaf area could be supported without rapid depletion of soil water. The water potential levels of the canyon trees may have been due to reduced root growth associated with shade-acclimation (2) that has resulted in more negative predawn water potentials in understory trees (1).
Conclusions
Reduced irradiance in the canyon environment may have resulted in low-light stress, as indicated by shade acclimation, but had the benefit of reducing tree transpiration during shaded periods. Tree transpiration was further reduced more than might be expected from lower radiation levels alone because shade acclimation resulted in greater stomatal sensitivity and closure at low humidities during the sunlit period. Consequently water stress arising from high transpiration rates and rapid soil-water depletion is less likely to be a problem for trees growing in urban canyons than those growing in full sun conditions. Soil conditions, however, are a very important consideration and water stress resulting from either confined or compacted soils would still be a concern.
- © 1993, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.