
Evergreen chapparal species in the foothills of California usually receive no rainfall for up to 6 months. In September, soil moisture in the root zone of these non-irrigated areas is probably less than 2 atmospheres depending on soil structure, and yet the species provide excellent green foliage cover. If there are several consecutive years of low winter and spring rains (this is drought in California), injury becomes apparent and trees and shrubs may die. In the more mesic regions of the United States many secondary diseases may be induced by drought episodes that occurred a year or two earlier (22). However, these disease-related symptoms are rarely observed in California's native species in their perennially droughty habitats.
Some chapparal species, e.g., toyon, ceanothus, and manzanita, make handsome landscape plants in many park, garden and freeway locations, but they are often irrigated and fertilized during establishment. This is intended to increase growth rate so that the plants make an impact on the landscape much more rapidly than would be the case with natural rainfall and soil fertility. The plants also develop larger shoot/root ratios than is the case in nature. The evapotranspirative (ETo) surface is larger than it should be for drought tolerance, and hence irrigation is required to maintain the established leaf surface area. In short, the plants are now acclimated to a high water environment and have lost their natural drought tolerance. Reduced landscape water use can be achieved once more by reducing irrigation and reacclimating the plants: that is the essence of the chapparalization of established landscapes. In previous work, completed in the 1970s, we found that considerable drought tolerance was gained by acclimating even mesic plants to a dry environment. A lowered shoot:root ratio was achieved (1) and other processes probably contributed to reduced water use, such as increased resistance to water vapor transport across leaves and lowered leaf water potential (3,9,10,11,12). Our results from 1971-76 and 1989-92, which will be reviewed below, suggest that 15 to 25% of ETo can be applied to most established shrubs and trees in the April to October interval with no loss of landscape function (2, 7, 8), providing that early spring soil moisture is adequate and the plants have been acclimatized by irrigation scheduling that does not exceed 25% ETo. At least a 50% reduction in vegetative growth is realized with this kind of irrigation regime, which by itself greatly reduces the cost of landscape maintenance. Owing to the reduced water demands, further savings can be gained by reduction of the size of irrigation pipes, valves, and meters required for any given system. We concluded that irrigation management rather than species modification was the key to reduced irrigation of established landscapes. The same management can be used during the establishment phase, but the cost will be reduced growth rate and delay in attaining the screening, shade or ornamental function.
Our work has not been applied perhaps because it is not sufficiently scientific, and because it ignores the notion that crop coefficients could be useful in determining irrigation practices. There is now a growing literature (14,15,16) addressing the importance of landscape coefficient (something akin to crop coefficient) in determining irrigation protocols. Use of crop coefficients has been deemed a better approach to the minimum irrigation problem in California and elsewhere even though these coefficients must vary according to site, climate, and irrigation strategy. That is, even if crop coefficients were useful, the research required to determine minimum irrigation for satisfactory appearance of plants for any site (landscape coefficient) is daunting. Intuition alone tells us that i n a mixed landscape the best that we can hope for is 1) to acclimatize a landscape to the maximum possible by correct management protocols, and 2) to specify an irrigation regime that satisfies the requirements of the least tolerant species.
We believe that certain simple guidelines can be used by way of irrigation scheduling to acclimatize a landscape. A key issue will be to insure high soil moisture in the spring. There is, unfortunately, a persistent belief that deep irrigations are preferable to more frequent, shallow irrigations. No data substantiate the preference. Frequent, low volume irrigations may improve plant performance using less water overall. In all cases irrigation frequency should vary inversely with effective rooting depth. Automated drip irrigation systems and soil moisture sensors provide the technology required to schedule irrigation and to irrigate at a pre-set level of soil moisture tension in the zone where most of the root system is active. The in situ technology for estimating when and whether established plants require irrigation is still being developed, but some methods, such as tracking evapotranspiration and estimating demand, are available and are used now.
Since our experience suggests that California landscapes may receive more waterthan required, and not always at the time required, objectives of future work will be 1) to determine irrigation timing and minimum water requirements for annual leaf renewal, which we believe to be crucial to plant longevity 2) to evaluate several species for appearance as a function of leaf area index and to develop quantitative methods for evaluation of plant appearance (looking for generalizations across species) 3) to measure water consumption of landscapes as afunction of some minimum leaf area index; to determine the impact of avoidance mechanisms on water consumption and finally 4) to evaluate soil moisture sensors for dependability in the 2-5% soil moisture range (less than - 1 atmosphere soil moisture tension for most soils).
Prior Experimental Work
Plant materials, sites, and methodology
From 1971 through 1974, hedgerow plantings of Ligustrum lucidum (glossy privet), Pittosporum tobira, Nerium oleander, Coprosma baueri, Xylosma congestum, Eugenia uniflora, Hedera (Algerian ivy), and Carpobrotus (ice plant) at experiment stations at Irvine (Orange County) and San Jose (Santa Clara County) were subjected to low irrigation regimes calculated as a fraction of reference evapotranspiration (ETo). ETo approximates the amount of water evaporated from a large field of an adequately irrigated, 4- to 6-inch-tall cool-season grass (14). The hedgerows and ground covers were established in San Jose and Irvine in 1965. Six years later, the plantings were subjected to three or four different irrigation regimes: (1)replacement ETo, (2)one or two irrigations beginning 30 days following the last significant rainfall (see figs.2 and 3), and (3)no irrigation. At both San Jose and Irvine, no significant rain fell after March 1974; hence, all water provided to plants was via irrigation. All irrigation treatments began with the soil profile wet to a depth of 4 feet, that is, they received one 3 inch irrigation in late March. Irrigation in the 1971-74 experiments, on the other hand, was by row and furrow with 3 to 4 inches applied at each irrigation. The volume of water applied was calculated from the rate of flow of water from the irrigation pipe and the duration of each irrigation. Further irrigation research at Irvine was begun in 1989 using oleander, ligustrum, escallonia, pittosporum and carissa. Chemical growth retardants have been used to determine whether performance of plants with low irrigation regimes is improved by growth retardation. Each species was planted in 1986, and hence well-established by 1989, according to the following plan: 2 irrigation blocks of 3 rows per block and 18-20 plants per row on 4 ft centers. For each species and irrigation block there were two border rows, 12 ft to either side of the test row. For each test row the two end plants were used as guard plants and between each growth retardant treatment there were two guard plants. These guard plants are essential since the lateral extent of the root systems is unchecked except by competition from neighboring plants. Each irrigation block was fitted with an automated drip irrigation system with soil moisture sensor set at a depth of 2ft in the ligustrum test row. The tensiometer was set at −20 cb for the ‘wet’ treatment. Based on experiments with eucalyptus plantations (15), we estimated that this setting insured approximately 0.75 ETo. Data recorded for March through July 1992 indicated that this was a correct estimate. The tensiometer was equipped with a magnetically activated switch that controlled a solenoid valve linked to a clock that controlled the on-off cycle of solenoid valves. Each irrigation cycle of the ‘dry’ treatment was set for one quarter that of the wet block (16 minutes maximum), with up to 8 cycles daily possible if the wet block tensiometer fell below −20 cb. The oleander, pittosporum, escallonia, and carissa plantings received the same irrigation regime as the ligustrum. The oleander and pittosporum canopies for the wet blocks were approximately the same densities (not extent) as that for ligustrum, whereas the canopies for escallonia and carissa were considerably smaller (Fig. 3 growth data).
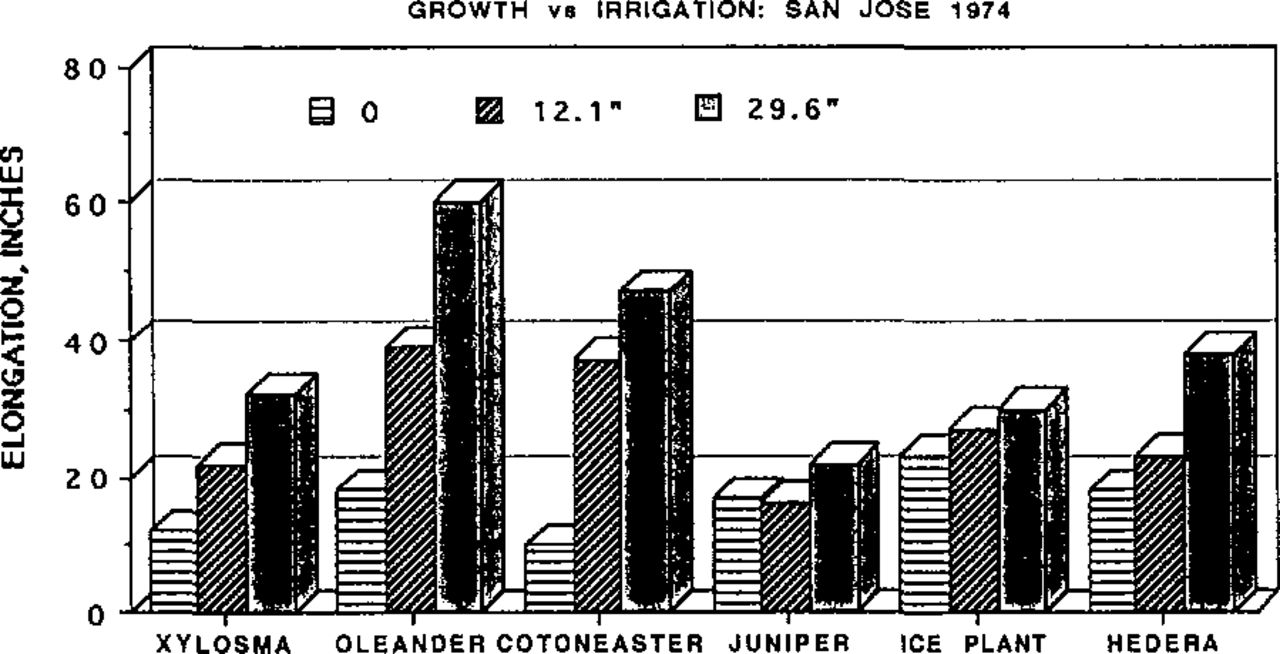
Growth of shrubs at San Jose as a function of irrigation amounts from the end of March through September 1974. The 12.1 in were applied in three irrigations in May, July and September; the 29.6 in were applied in eight irrigations at 2-week intervals beginning in April. Rainfall for July 1973 through June 74 was 15.2 inches.

Fig. 3. Negative print of aerial infrared photo of San Jose plantings taken in May, 1976, the 5th year following irrigation treatments. From top to bottom the species are: Pinus radiata, Juniperis chinensis, Xylosma congestum, Nerium oleander, Cotoneaster parneyi, and ice plant. The 3 blocks from left to right are the irrigation treatments: 0,12.1, and 29.6 inches of water applied annually in the April to November.
Evapotranspiration at San Jose and Irvine
Ten-year average ETo values were used for scheduling irrigation for 1973-74. From March through September, the periods for which growth and appearance data were recorded, the cumulative ETo was 30.4 inches at the South Coast Field Station and 29 inches at San Jose.
Results
Growth and appearance of plants
In 1974, the third year after the beginning of the irrigation treatments, by which time the plants in the zero- and low-irrigation regimes were “hardened”,growth data were recorded (Figs1, 2).

Growth of shrubs at Irvine as a function of irrigation water received from March through September 1974. No bar appears for “0 inches irrigation” because no growth occurred under this condition. Each irrigation delivered 3.7 inches. Rainfall during the winter of 1973-74 was 10.9 inches. All but the nonirrigated plants retained an acceptable appearance for landscape purposes.
Vegetative growth was reduced substantially in the low-irrigation treatments, so that the shrubs would not have required pruning (and, hence, no vegetation removal). Less than 4 inches of irrigation water, approximately one-eighth of the ETo, was required to maintain shrubs in a healthy condition. All species retained an appearance suitable for most landscape purposes. Only the non-irrigated shrubs at Irvine suffered significant leaf loss; their appearance was not acceptable. Yet the single irrigation in May, supplying 3.7 inches of water, or 12.5% ETo, was adequate to maintain plants in excellent condition. At San Jose all species in the non-irrigated treatments were in excellent condition as revealed by the infrared photographs taken in 1976, more than 5 years after the irrigation treatments were begun (Fig. 3).
Note that rainfall at San Jose was 5 inches greater than at Irvine during the winter of 1973-74 and ETo somewhat lower than at Irvine. During a period of below average winter rainfall (less than 10 inches), 1991 results show clearly that growth is reduced approximately 50% by irrigation set for 0.25 ETo (Fig. 4). So far in 1992, with rainfall in excess of 7 inches during the period of observation, the 0.25 ETo block appears to have grown significantly less than the ETo block.

Growth of shrubs at Irvine, 1991, as a function of irrigation regime and growth retardant application. Flur equal to flurprimidol, Clippings were weighed in October 1991,11 months after pruning. Irrigation was automatic. A tensiometer with a magnetic switch set at −20 centibars controlled the irrigation valves. The controlling tensiometer was placed in the ligustrum block under an emitter at a two feet depth. Irrigation equal to ETo was delivered with the tensiometer set at − 20 cbars. For each irrigation cycle, the 0.25 ETo shrubs were irrigated for a period one quarter as long as the ETo block. Precipitation for September 1990 through October 1991 was 14.07 inches, with 5.86 inches falling in March 1991.
Leaf area of the zero- and low-irrigated plants was significantly reduced during the periods of adaptation. Before the irrigation experiments were begun, all plants were grown at near replacement ETo irrigation, thus supporting maximum growth rates. The plants developed lush canopies, which required substantial pruning annually (growth data for the 29.6 inch irrigation regime Fig. 1, 2 and the ETo data of Fig. 4).
Growth retardants
The 1990 test at Irvine used growth retardants (flurprimidol, initially) to induce more rapid drought hardening of plants in low irrigation regimes. The results suggested that indeed these compounds may have value for the intended purpose. Regardless of irrigation regime, soil-applied flurprimidol reduced the rate of stem elongation producing more compact plants (Fig.4). Although growth retardants may increase drought hardiness by subtle effects on plant metabolism, they quite obviously reduce leaf surface area and this is undoubtedly the major reason that they reduce evapotranspiration. We have found that in factthe 0.25 ETo, growth retardant-treated shrubs had a reduced canopy that appeared denser and more complete (less gaps through which to see the barren branches) because they were so compact. Their appearance was improved and growth was reduced.
Leaf temperature
Leaf temperature is used to measure moisture stress and irrigation requirements in some landscape and crop species, but has not been used to control irrigation where minimum growth and satisfactory plant appearance are desired. We used a remote-sensing, thermalimaging system to measure leaf, air and soil temperatures. Plants at San Jose that were not irrigated were generally warmer than those that were irrigated (Fig.5). Ice plants were up to 25°F warmer than the surrounding air when air movement was minimal, less than 5miles per hour. With wind velocities above 5mph, leaf temperatures in the nonirrigated ice plants were close to ambient air temperature. Hence, the leaf temperature method of measuring stress would not be useful in most areas of California where wind velocities are frequently greater than 5mph, unless the relationship between wind velocity and leaf temperature were known with some precision. Xylosma foliage, further above the ground, with thinner leaves that dissipate heat by conduction and convection more rapidly than those of ice plant, were only 10°F warmer than the surrounding air when air movement was less than 5 mph.

Leaf-air temperature differentials for San Jose Xylosma and ice plant as a function of irrigation regime. Temperatures were measured July 26, 1974, using infrared imaging system. Air temp. =76°F.
Soil moisture, rooting depth, sensors and sensor placement
The well-irrigated shrubs at Irvine and Santa Ana had an average rooting depth of 4 to 6 ft. Soil profiles were wet down to a least 4 ft at the beginning of the irrigation trials in April. Although no quantitative estimates were made, the bulk of the root systems in the nonirrigated and infrequently irrigated blocks was in the upper 2 to 3 ft. Below this level, roots were not apparent (fine tertiary branch roots would have escaped attention by the coarse retrieval methods used). Soil moisture was extracted from at least the upper 3 ft in all irrigation treatments at Irvine and San Jose (Figs. 6 and 7) and was below 5% in the nonirrigated plots. Tensiometer readings for the monthly irrigated plots were −40 to −60 centibars. In nonirrigated blocks, readings could not be determined because they fell below −80 centibars, a region where tensiometer accuracy is questionable. There is a need for sensors that can function reliably in the 2-5% soil moisture range. In a drip irrigation system, with emitters irrigating only a portion of the roots, sensor placement is critical. Experiments with oleander and pittosporum shrubs suggest that a capacitance-type sensor placed at the one-foot level of the root zone, about 1 ft from an emitter, is an effective system regulator, but we have not yet determined whether it can be adjusted to the low soil moisture desired. Our attempts to calibrate the sensor in this range, using glycerol/water solutions in the 1 to 5% range, have so far proved unsatisfactory.

Soil moisture as a function of depth and irrigation regime at Irvine. Cotoneaster plants extracted water from at least a 3-ft depth in the nonirrigated block (these shrubs suffered leaf damage). Measurements were taken in October, 1974; rainfall from July 1973 through June 1974 was 10.9 inches.
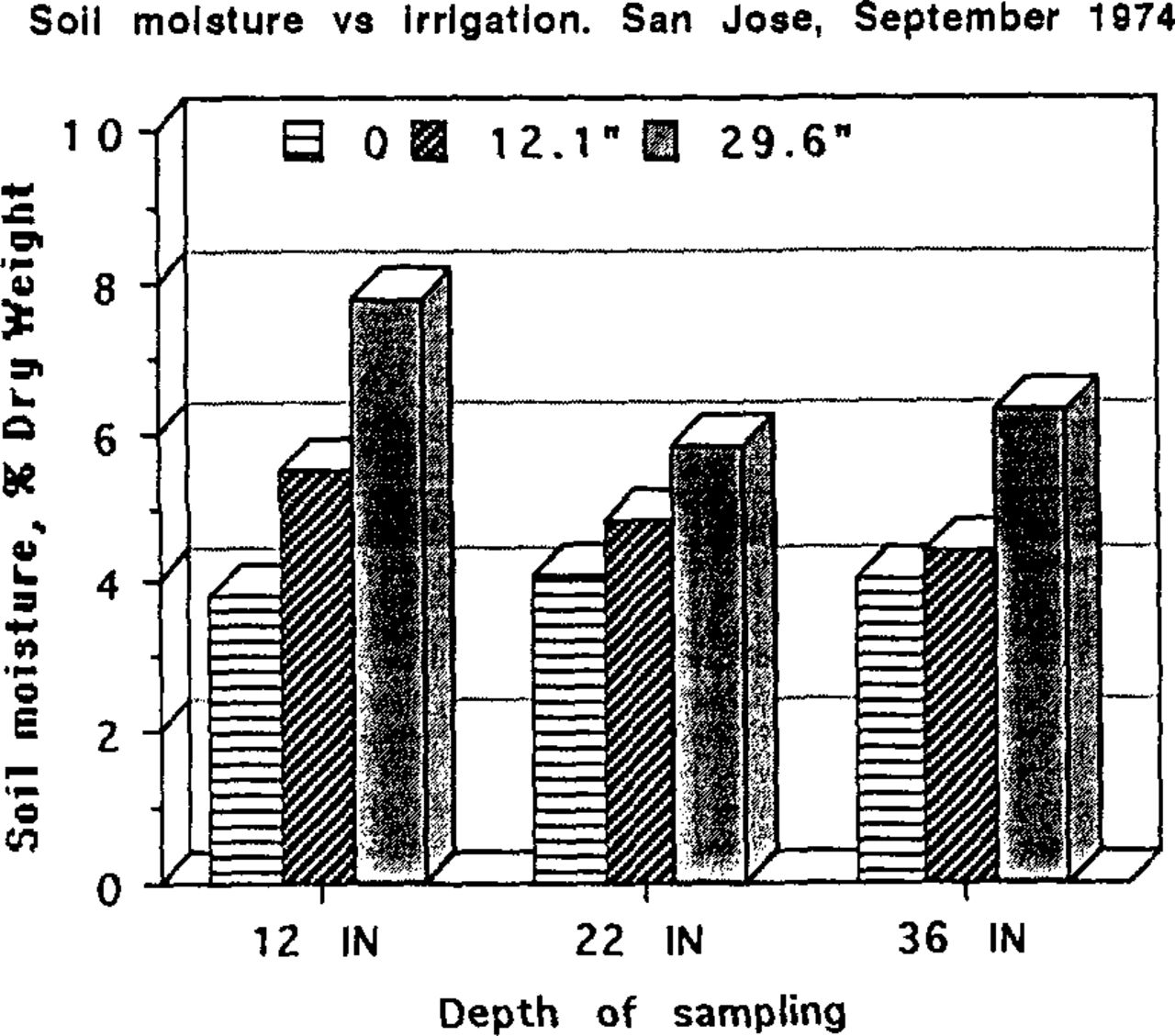
Soil moisture as a function of depth and irrigation regime at San Jose. Soil samples were taken from below Xylosma plantings, at 12,22 and 36 inches, on September 16,1974.
Conclusions
Irrigation duration should be related to rooting depth
More frequent irrigations of shorter duration (than used on our test blocks) are recommended if the effective rooting depth of the plants is less than 3 ft. Minimally irrigated plants have reduced leaf surface areas, reduced total plant photosynthesis, and therefore less metabolites for root system development. That is, in a minimally irrigated plant, the absolute size of the root system is reduced, even though the root:shoot ratio is larger. Rooting depths may be less than 3 ft in heavy clay or compacted soils due to restricted oxygen supply. In this situation deep irrigation is useless.
Leaf renewal, hardening and spring irrigation
Many species do not develop cold hardiness unless they can photosynthesize during the hardening period (19,20). We found that during the initial 4 weeks of low, non-freezing temperature exposure, eucalyptus, tamarisk and acacia plants did not develop freeze tolerance if temperatures were too low during the day, or if they were drought or salt stressed (Sachs, Booth, Cartwright, and Lee 1991; unpublished). Freeze hardening is clearly an active process and we believe that this is probably true for drought hardening as well. Thus, a significant factor in drought hardening is soil moisture in the spring sufficient to sustain normal metabolic functions. These functions are required for hardening as well as canopy renewal. In many ‘evergreen’ shrubs and trees, leaves age and die annually, often in the spring of the 2nd year. Hence, even in drought conditions, with the eventual goal of reducing the plant canopy, some minimal leaf renewal in the spring is a key process if a plant is to retain satisfactory landscape appearance. If the winter is extremely dry, a landscape should be irrigated in late March or April to insure that plants can renew their canopy and adapt to the following drought period. In California the ‘March’ miracle of 1991 (the nearly 6 inches of rain at the end of March) may have served that purpose.
All irrigation studies at Irvine and San Jose were begun in April of each year with the upper 3-4 foot soil profile at field capacity. The plants had ample water in the spring when leaf initiation and canopy renewal began, and the new leaves could photosynthesize when spring temperatures were near optimum. In March 1992, one block of ligustrum, oleander, and pittosporum plantings at Irvine received 3.6 inches of rain, less than 0.25 inch of irrigation, with little decline in appearance.
Current photosynthesis permits gradual adaptation to reductions in soil moisture. Plants continue photosynthesizing until leaf water potential is lowered to cause near complete stomatal closure (by which time leaf renewal and adaptation to water stress would be complete). Hardening is an annual process, beginning each April with a new canopy, to meet the eventual stress of the long April through November drought period. This is the annual routine for chapparal. For this reason we believe that an appropriate management strategy is a) to make certain that soil moisture is high, say - 20 cbars, down to 3 ft in April, and b) to cut irrigation until soil moistures fall to 3 to 5% near the end of summer.
Can these results be extended to other climatic regions?
The thesis that acclimatization through controlled, minimum irrigation is the best way to prepare plants for drought and avoid drought-related injury, may work well for California and other Mediterranean-type climates where precipitation is limited to the winter-spring months and growing season rainfall is virtually nil (certainly well below ETo). Yet it is impractical in mesic environments where rainfall is usually plentiful during the growing season and, hence, natural acclimatization does not occur. In high rainfall areas, growth retardants that reduce shoot:root ratios should be helpful to help plants avoid midsummer droughts. Low soil moistures in the spring, during the period of leaf renewal, may prove to be the most serious aspect of drought and at least for the present may be prevented only by supplemental irrigation or severe pruning of the canopy to reduce the shoot:root ratio. Lindsey‘s work on water use for several species in New York State suggest that plants use less than 40% pan evaporation under optimal conditions for growth (21, Fig. 8). Hence, minimum irrigation during drought periods, where only leaf renewal or tree survival need be insured, may be feasible.

Comparison of water lost from trees and pan evaporation (PAN)on Aug.22, 1991 in New York. SOPH ≡ Sophora japonica, FRAX = Fraxinus americana, AMELANCH = Amelanchier, TIL = Tilia americana (from Lindsey (21).
- © 1993, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.