
Abstract
Biotechnology, which has made significant contributions to the improvement programs of agronomic crops, offers the opportunities to enhance forestry research and accelerate tree improvement. Forest biologists and tree breeders are turning their attentions to these biotechnologies, which enable them to overcome barriers and can be integrated into conventional breeding methods leading to more rapid progress in tree breeding. Plant biotechnology currently comprises a range of activities, such as vegetative propagation and tissue culture, genome analysis and gene cloning, DNA recombination and gene transfer, and DNA-based selection. Although application of biotechnology in forest trees and ornamental woody plants is just in its infancy, micropropagation is rapidly becoming a standard tool for tree improvement. In addition, emerging success and practical application have become visible in genetic transformation. These initial achievements have already proven that biotechnology will make inestimable impact on tree improvement.
It is appropriate to ask, at the beginning of this review, what biotechnology means and how it relates to conventional biology. The word is easy to use but difficult to define. Biotechnology is a collection of new technologies applied to biological systems rather than a separate academic discipline. Thus, it may be defined as the manipulation of biological systems (organisms or biological processes) via modern technologies to solve practical problems in agriculture, medicine and industry. Plant biotechnology may include clonal propagation via tissue culture, protoplast fusion, gene cloning, DNA recombination and genetic engineering, mutant induction and in vitro genetic selection. Molecular-marker-aided genome analysis and gene mapping are also sometimes included in plant biotechnology, as are aspects of agriculture, horticulture, and forestry where genetic analysis and gene identification are used in plant breeding programs.
The large size, long regeneration cycle, and sporadic seed production of trees causes numerous problems in tree improvement programs, such as the time in years required to reach reproductive maturity, low efficiency in producing new and improved germplasms, space limitations, time consumption and great cost. Over the last several decades, however, forest genetics and tree improvement research has shown great advances. Traditionally, tree breeders have employed conventional methods, i.e., breeding and selection to make genetic combinations for creating new and improved forest stocks. Unfortunately, these traditional methods for introduction of desirable traits from one genotype to another are tedious, costly, time consuming, and not always successful. Even when potentially useful improved lines or hybrids are developed, many years of field testing are needed to prove their usefulness. However, the newly developing biotechnologies, such as tissue culture, genome analysis and gene identification, DNA recombination and genetic transformation, and RFLP/RAPD markers, provide the opportunities to solve the above problems and to accelerate tree improvement. No doubt, trees are the crops for which biotechnology offers unique potential benefit.
The recent advances in plant biotechnology and the achievements of research in woody plants have shown that the potential for application of genetic engineering to forest trees and ornamental crops is great. In recent years, we have concentrated our efforts on developing: 1) micropropagation methods for clonally propagating superior conifers, 2) systems for gene transfer and regeneration of transgenic trees, and 3) initiation of studies on genome mapping of abiotic stress traits in loblolly and shortleaf pine using RFLP and RAPD technology. In this paper, we will first review some key biotechnologies and their advances. Then, we will discuss the possibility of extending the developed technologies to other forest tree species and ornamental woody plants and assess the potential impact of plant biotechnology on tree improvement, management and forest productivity.
Tissue Culture in Woody Plants
Forest trees and ornamental woody plants are important components of today's global society. Present global problems with increasing human populations and diminishing natural resouces demand the development of strategies for improving tree species and for growing more and improved trees on a decreasing land base (26). The potential of plant tissue culture includes: 1) enhanced production of natural products, 2) rapid clonal multiplication of select genotypes, 3) production of disease-free plants, 4) germplasm preservation, and 5) genetic manipulation. In the last decade, much progress has been made in woody plant tissue culture (2,80). In this section, we will briefly discuss the practical applications of tissue culture in improvement of forest and ornamental trees.
Micropropagation
This term is defined as an asexual propagation process using artificial techniques. Early vegetative propagation methods included rooting of cuttings, grafting and budding, which are standard methods in clonal forestry programs and in the horticulture industry. As in vitro technology has been developed, micropropagation has become a more effective tool for both basic research and commercial practices for some tree species. Since the pioneer regeneration work reported in Populus tremuloides and in Pinus palustris via organogenesis and in Ulmus americana via somatic embryogenesis in the early to the mid 1970s, micropropagation techniques have been developed and refined for many woody plant species, including both angiosperms and gymnosperms. Today, successful lab-scale protocols are well developed for about 70 hardwood and 30 softwood species (86).
Regeneration of plants via in vitro micropropagation systems can be achieved in various ways, such as promotion of axillary bud break, induction of adventitious buds, and development of somatic embryogenesis. In most cases, the simplest method for clonal propagation is to place sterilized shoot tips or axillary buds onto a culture medium to induce formation of multiple buds (Fig. 1). This method has been successfully employed for a variety of woody plant species, including a recalcitrant hardwood, Quercus robur (69). Mass micropropagation via adventitious bud production from juvenile tissues is another standard method for many plant species (2,20). However, the ease of adventitious bud initiation is strongly dependent on tree species. We have developed a reliable system for plant regeneration via adventitious bud initiation in Lar/xspecies (51). With a single pass throughout our culture system (Fig. 2), at least 70 new adventitious buds/shoots can be multiplied from each initial larch seedling or juvenile explant. Juvenility of plant materials is a key factor in vegetative propagation because the regeneration ability of woody plants decreases dramatically as they approach maturity. Nevertheless, there are a few reports of the successful induction of adventitious buds and establishment of plantlets from mature trees, such as in Sequoia sempervirens, Larix decidua, Picea abies, and Fraxinus ornus (4). On the other hand, rejuvenation can be obtained in some hardwood species by cutting or by inducing sprouting with hormone treatment.

Adventitious buds (A) developing from meristematic tissues in the apex of a Larix decidua seedling hypocotyl (A) and bud cluster (B) proliferating from existing bud on the upper branch of 4-year-old larch tree in vitro (x 8).
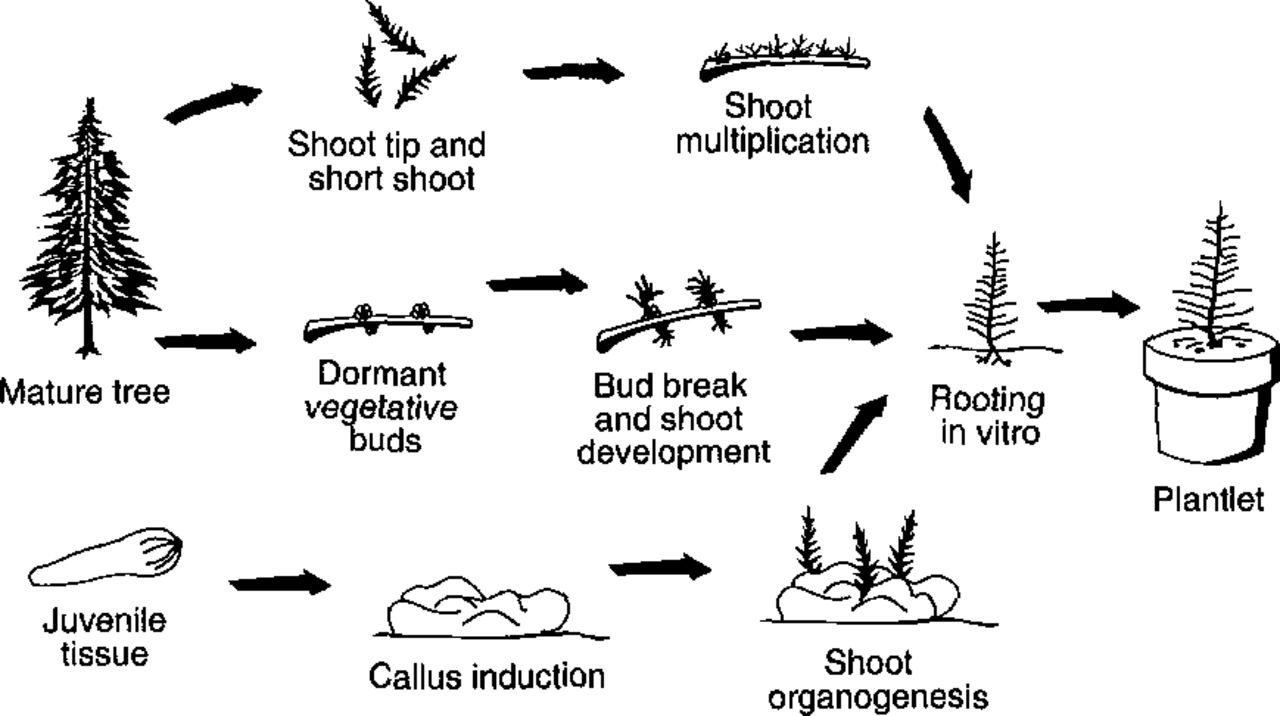
Developed processes for Larix micropropagation.
More advanced technologies for plant regeneration using tissue culture rely on development of plantlets from callus, cell and protoplast cultures or somatic embryogenesis. Such systems are based on the concept of the “totipotency” of living cells, that is, that each cell possesses all the genetic information leading to the development of an intact plant under favorable conditions. These regeneration systems are generally more difficult with woody plant species than the previously mentioned micropropagation systems. But their potential application is unmeasured in clonal propagation and genetic manipulation of trees. In the past decade, some major breakthroughs have been made in tree somatic embryogenesis as well as protoplast culture (2,20,25,35,39,56,74). In spite of these advances in laboratory research, much additional research is needed before these systems can be applied practically or transferred to commercial operation.
Regeneration of whole plants is the final step in the process of clonal propagation via tissue culture systems, and it is the critical step to determine whether or not the system is successful. Of tree species, gymnosperms such as mature pines, spruces and firs and some agiosperms such as oaks, maples, beeches, and walnuts continue to be difficult to regenerate in vitro (39). Success in tree regeneration greatly depends on the choice of tissue source, age of the tree, and species or genotype chosen. Whatever system is chosen for tissue culture of a given species, the ability to regenerate intact plants is essential. Finally, trees regenerated from a propagation system must perform satisfactorily in field tests to prove their practical value in tree improvement programs.
In general, plants regenerated by vegetative propagation are genetically uniform. Once an elite clone is identified, phenotypically uniform trees with its desired characteristics can be produced in mass in a short time period. The potential of mass propagation from tissue culture is great and multiplication of clones can continue indefinitely and exponentially with continuous in vitro culture. It has been reported that more than 60,000 apple shoots were produced from a single shoot tip in eight months (49). Aitken-Christie et al. (3) reported that 260,000 radiata pine (Pinus radiata) trees could be produced from one seed in 2.5 years using a meristematic tissue system.
Use of somaclonal variation
Genetic variation, known as somaclonal variation, has been observed in plants regenerated from tissue culture, particularly in cultures of callus, cells, orprotoplasts. This variation may result from chromosome changes, nuclear DNA changes or organellar DNA changes at certain stages during culture, such as DNA replication, mitosis or cell division. Somaclonal variation represents a new source of genetic variability; therefore, it constitutes another tool for plant breeders (54,89). Exposure of in vitro cultures to mutagenic agents ortostress conditions can increase the number of somaclonal variants. Using this system, successful selection of mutants with various desired characteristics has been reported in some plant species, such as increased herbicide tolerance in hybrid poplar and other species (60,13), disease resistance in Larix and other species (24,75), and heavy metal and salt tolerances in several species (61,77) as well as multi-gene agronomic characters (62). Serres et al. (79) reported somaclonal hybrid Populus variants that were dwarf but fast-growing and with color-changed leaves. Thus, selection of somaclonal variants is particularly useful in the creation of new ornamental characteristics. The problems with use of this technique are: 1) there is a low frequency of variants, so a large-scale experiment is needed, 2) most variants are not desirable (useful), and 3) many of the variants may be epigenetic in nature, and not true variants. In some cases, the resultant variants are due to stable mutations with sexual transmission of the traits to progeny (13,79) and thus a useful source of variation.
Protoplast fusion and somatic hybridization
Somatic cell hybridization via protoplast fusion is another alternative for plant improvement. In this process of parasexual hybridization, two protoplasts (with their cell walls removed by enzymes) are fused in culture medium, then the resultant hybrid cells are regenerated into whole plants in vitro. This system overcomes some restrictions in sexual reproduction, such as incompatibility, sterility and other problems, and is especially useful to produce interspecific or intergeneric hybrids. For example, resistance to Dutch elm disease from Asian elm (Ulmus parvifolia) has not been transfered to American elm (U. americana) with traditional breeding methods. Protoplast fusion could be an alternative way to achieve this gene transfer since the plant regeneration system from protoplasts is available in Ulmus hybrids (81). An excellent example of alien chromosome transfer through protoplast fusion was published by Gupta et al, (38). In their experiment, albino Datura protoplasts were fused with normal Physalis protoplasts. Some such achievements have been made in breeding programs of agricultural crops. At present, relatively few woody plants have been regenerated from protoplasts (56). In order to facilitate the use of protoplasts in tree breeding, efficient and reliable methods must be developed for regeneration of trees from protoplasts or cell cultures.
Disease screening and virus elimination
Tissue culture provides the opportunity to develop disease-free plants through in vitro diseasescreening and pathogen-eliminating strategies. Many tree diseases are transmitted by seeds or vegetative propagules from one generation to the next. However, it has been found that the active apices of shoots and roots are frequently free of pathogens, particularly viruses. One can dissect such meristems from infected plants, culture and regenerate new plants from these cultures, and subsequently obtain virus-free plants. This culture system can also be used to obtain disease-free plants from stocks systemically infected with mycoplasma, fungi and bacteria (90). Therefore, meristem cultures are of particular value to urban forestry nurseries and the horticulture industries. Micropropagation systems can also be useful for testing the response of trees to pathogens. In vitro larch scleroderris canker screening appears useful for selecting disease-resistant clones (1). Hubbes and Ho (47) conducted studies on pathogen genetics and on the mechanisms of elm resistance to the pathogenic fungus, Ophiostoma ulmi, employing tissue culture. It should be possible to screen elm genotypes in vitro for variability in response to the toxin and to utilize somaclonal variation in response to the toxin.
Germplasm preservation
Storage of genetic materials has become increasingly important for developing new cultivars as well as preserving heirloom varieties and rare or endangered species (72). Two tissue culture approaches have been developed for germplasm preservation, in vitro conservation and cryopreservation. These advances contribute to the establishment of an “in vitro genebank” of plant germplasms. In vitro conservation maintains plant cultures at low temperature and low light intensity. This in vitro long-term storage of plant cells, tissues and organs is often favored because it can greatly reduce the labor and space requirements of traditional storage methods. For example, meristem cultures and shoot-tip cultures of vanilla and Musa can be maintained on filter paper bridging in liquid medium for 18 months (48).
Many genotypes and even species of trees are being lost due to enivronmental or disease problems. For instance, some populations of Larix decidua are being threatened by air pollution, and American chestnut and American elm are being ravaged by disease (51). Cryopreservation, freezing in vitro cultures with liquid nitrogen and storing at −30 to −196°C for years, would be an appropriate method to preserve these valuable germplasms for the relatively long-term. In addition, cryopreservation is a secure and reliable conservation method with a high level of genetic stability (37). Survival of such stored cultures has been demonstrated in various types of plant cells and tissues (33), including a number of forest and horticultural species, among them Rubus and Pyrus (72), Acer pseudoplatanus, Populus, and Ulmus americana (50).
Bridging the development of technologies with their practical application
Micropropagation techniques are being rapidly advanced in research laboratories. At present, successful tissue culture systems have been developed for many woody species. There is no doubt that tissue culture technology has had a great impact in tree improvement programs. Clonal propagation and tissue culture could become a standard method for regeneration of long-living trees. However, not many of these newly developed techniques have been yettransferred to operational practice. Zobel (97) reported a lapse between the development of a technique and its operational use in forest vegetative propagation. Tissue culture specialists frequently concentrate on developing technology to produce plantlets in laboratory conditions but generally ignore the refinement of their protocols for operational field scale conditions. On the other hand, field foresters or tree breeders often do not have an adequate working knowledge in tissue culture. Thus, with few exceptions, successful tissue culture technologies have not yet been used for practical commercial purpose. In fact, this problem can be easily resolved by cooperative research programs. Although interest in clonal propagation and tissue culture in forestry is apparently high, the research funding to develop these systems for practical programs remains limited.
Gene Transfer and Genetic Engineering of Trees
The formation of new combinations of heritable material can be achieved by traditional breeding techniques and practices, or by genetic engineering. Genetic engineering of plants usually implies direct genetic manipulation of plant cells at the cellular or molecular level, adding new genes to the plant’s genome or replacing certain genes with other genes from other sources using recombinant DNA technology. The modern techniques developed for genetic engineering include vector-mediated systems based on Agrobacterium plasmids or viral vectors, and non-vector systems (direct gene transfer), such as biolistics (particle bombardment), microinjection, electroporation (electrical pulse treatment), chemical poration (polyethyleneglycol-mediated osmotic DNA insertion), liposome fusion, and protoplast fusion.
Development of gene transfer technology
Genetic engineering depends upon both the development of gene transfer systems and the availability of valuable genes. Rapid progress has been made in developing technologies for introducing genes into plant cells in the past several years (70).
Agrobacterium-medìated Gene Transfer
The first practical, most successful and most widely used method for genetic engineering of plants relies on Agrobacterium-mediated genetic transformation systems. Agrobacteria are plant pathogenic organisms that cause tumoric diseases on infected host plants. The exploration of this gene vector system has developed from an understanding of the molecular basis of its pathogenesis. Agrobacterium tumefaciens and A. rhizo-genes harbor an additional (nonchromosomal) genetic component, called the Ti(fortumor-inducing) or Ri (for root-inducing) plasmid. This virulent plasmid (about 200 kb) is the disease-causing agent because it carries the phytohormone biosynthesis genes, which are located on a T-DNA (for transferred DNA) region (17,40). During infection, Agrobacterium inserts its T-DNA into plant cells, and these inserted DNA fragments are then integrated into the plant chromosomes. These transferred hormone genes replicate and express along with the plant’s DNA so that the elevated levels of phytohormones cause the tumorous growth with abnormal patterns, such as crown galls or prolific root masses. However, the genes causing tumorous phenotype can be removed by deleting them from the T-DNA without loss of DNA transfer and integration functions.
In order to develop the Agrobacterium plasmid as a gene vector for genetic engineering of plants, researchers have genetically engineered the T-DNA by replacing the oncogenic genes (tumor causing) with useful genes. In this way, the first genetically engineered plants expressing functional foreign genes were produced using tobacco less than ten years ago (21,41). Thereafter, Ti and Ri plasmids, the natural gene vectors, have become an important tool for genetic engineering of plants. They are now routinely used in many research laboratories and commercial companies around the world. Researchers have also applied this system to tree improvement. Successful gene transfer by A. tumefaciens vectors was first reported in hybrid poplar, a woody angiosperm (30,67). Furthermore, a simple and reliable system was developed for transformation and regeneration of a gymnosperm species based on the use of an Ri plasmid vector of A. rhizogenes (Figure 3). Thus, the first transgenic conifer plants were produced in European larch (Fig. 4) by such a system (43,45). DNA transfer using the Agrobacterium vector has been demonstrated in many woody plants but without subsequent plant regeneration. A. tumefaciens and A. rhizogenes have proven to be excellent vector systems for the production of transgenic plants (32,34). Agrobacterium-mediated genetic transformation systems have been successfully applied to many plant species including several woody plants (Table 1). Use of the Agrobacterium system is restricted by its host-range, since some plant species are not susceptible to infection with Agrobacterium. Other types of biological vector systems are plant DNA and RNA viruses. It has been possible to introduce foreign genes into plant cells using viral vectors (10). However, forsometechnical reasons, little practical application has been made of these viral vector systems.
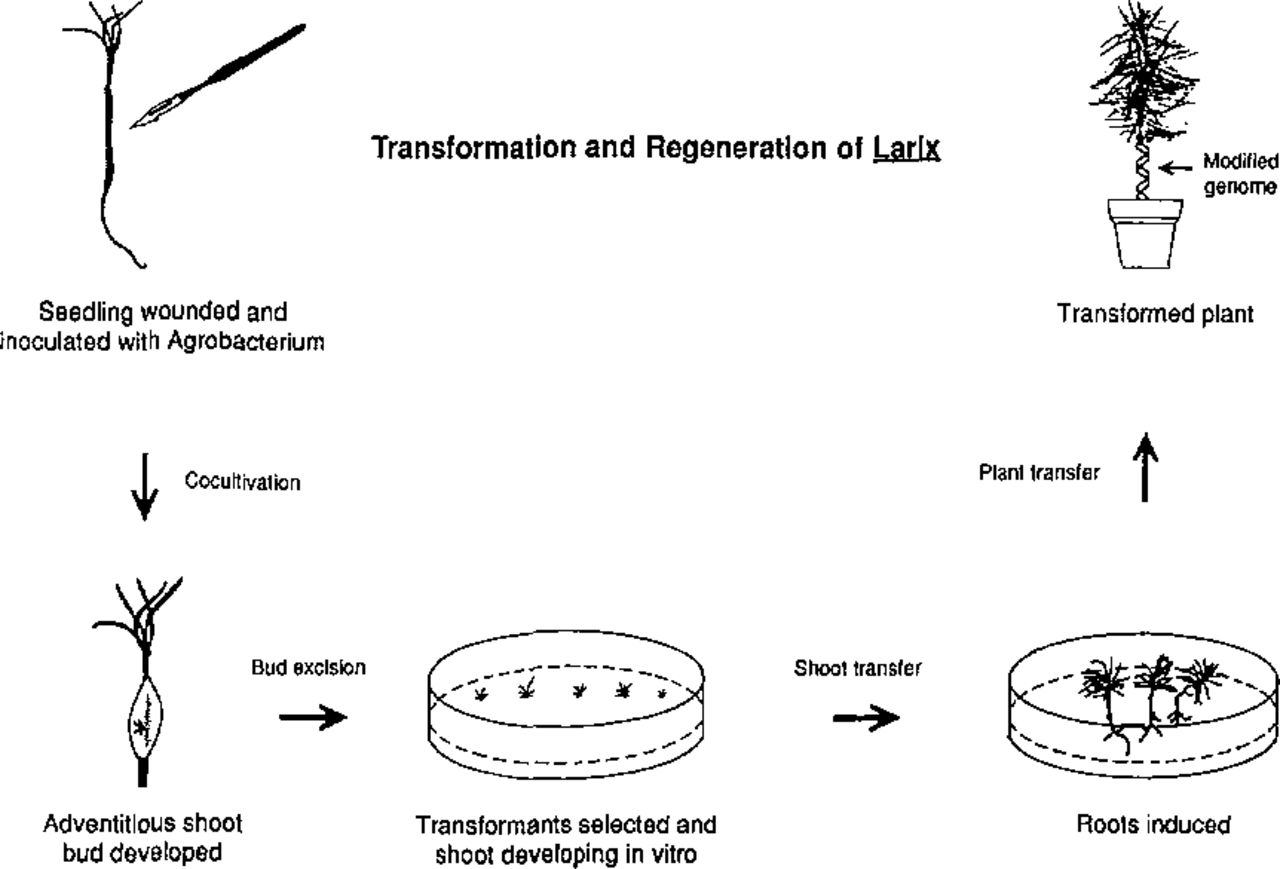
A simple and effective system developed for genetic transformation and regeneration of transgenic plants in European larch using an Agrobacterium rhizogene-mediated gene transfer vector.

Transgenic plantlet of Larix decidua with well-developed shoot and roots.
Woody species in which transgenic plants have been produced.
The authors have developed a simple and effective system for genetic transformation and regeneration of transgenic plants for an economically important conifer, Larix decidua (Fig. 3) (43). Transgenic plants have been reproducibly developed which express for two agronomically important foreign genes, the aroA gene which renders plants tolerant to the herbicide glyphosate, and the Bt(Bacillus thuringiensis) insect toxin gene that protects plants from harmful insects, such as Lepidopterans (44). This first achievement in production of a genetically engineered larch represents an important milestone in genetic engineering of gymnosperms and may serve as a model system for genetic engineering in other conifers.
More recently, Huang and Tauer (46) demonstrated integrative transformation of the recalcitrant species Pinus taeda, P. echinata and a P. taedax P. elliottii hybrid. This progress represents a significant advance in genetic transformation of pine species with the Agrobacterium-mediated transformation systems.
Direct Gene Transfer
In addition to the Agrobacterium-mediated gene transfer systems, a great variety of alternative approaches have been explored for direct gene transfer. Electroporation has recently emerged as a predominant method for protoplast transformation. This method involves the application of a high-voltage electrical pulse to a solution containing a mixture of protoplasts and foreign DNA. This process facilitates the movement of DNA molecules into plant cells through transient openings in the plasmolemma (protoplast membrane) caused by the electrical pulse treatment (31). For protoplasts, it has been one of several techniques for routine and efficient gene transfer in plants. Microinjection and macroinjection, the injecting of DNA into single plant cells, successfully established for transformation of animal cells, are being adapted for transformation of plant cells (18). These micromanipulation techniques have the capacity to deliver foreign DNA into intact regenerable cells and may avoid the inherent difficulties of plant regeneration from protoplast and cell cultures. Trangenic plants have already been produced using these techniques (22,64).
Other vectorless gene transfer systems have also been explored for plant genetic manipulation, such as liposome-mediated DNA delivery, polyethyleneglycol-mediated direct DNA uptake, and pollen-mediated gene transfer (70). Although most of the above mentioned non-vector methods need further development for practical application, they show promise as plant genetic engineering systems. Some of these non-vector techniques have been also successfully applied to transformation of tree species (6,14,95). These few examples demonstrate their potential for introduction of foreign DNA into woody plant cells and tissues. Indeed, for some species, one of the direct gene transfer techniques is the method of choice for achieving gene transfer. However, these systems are still under study because of many limiting factors. For instance, incorporation of foreign DNA into a plant genome is sometimes unstable, and cell/protoplast culture systems are required, but regeneration of whole plants after transformation is not yet possible.
The development of the gene gun (particle bombardment or biolistics) is another approach for foreign DNA insertion that has recently received considerable attention. This physical method involves acceleration of DNA-coated heavy microparticles (microprojectiles) for directly delivering foreign DNA into plant cells and tissues (76,55). Over the past few years, this method has evolved into a useful tool for direct gene transfer to every type of living cell and intact tissue from plants, animals and microorganisms (52). The early efforts demonstrated gene transfer and transient expression of marker genes in onion cells (76). Later, stable transformation was achieved in an agriculturally important crop, soybean (55). Since then, this method has been applied to studies of gene transfer and expression in many plant species. Successful gene transfer with particle bombardment has also been reported in a few of tree species, including Pinus taeda (82), Picea glauca (29), and Populus spp. (58). The biolistics system has advantages and potential for general applicability: it is easy to handle, and it delivers DNA into many cells each shot, targeting every type of cell and even intact tissues. But integrative transformation events from biolistic experiments are of relatively low frequency and the technique is inefficient in yielding stable transformation (70).
To date, foreign genes can be transferred to plant cells and tissues by various DNA transfer systems. Subsequently, transgenic plants can be recovered via a variety of regeneration systems. Successful systems have been developed for a wide range of plant species, including some woody angiosperms and few gymnosperms. However, the ease with which production of genetically engineered plants can be accomplished varies between species. Therefore, a suitable system needs to be chosen or developed for each species.
Traits of interest for tree improvement
The success of producing genetically engineered plants relies on not only the development of a gene transfer system but also on the availability of desirable genes. Genetic engineering, like conventional tree breeding, will focus on using superior genes or desired traits to genetically improve trees. Although few useful genes have yet been cloned from trees (11), advances in gene cloning and recombinant DNA technology open the door for forest biologists. Some beneficial genes have already been isolated from other organisms and successfully transfered to target plant species. For example, single gene traits that help plants resist harmful insects, pathogens, and herbicides are desirable targets to incorporate into tree breeding programs, and some of this work has already been accomplished.
Traits for Tree Management
Genes detoxifying several selective herbicides are available, such as the aroA gene or EPSPgene for glyphosate (Roundup), the bar gene for phosphinothricin (Basta), the bxn gene for bromoxynil, and the ALS gene for sulphonylurea and imidazolinone herbicides (78). Transgenic trees carrying the aroA gene have been produced in Populus(30) and Larix decidua (42). Development of trees that are tolerant to herbicides is an important component in weed control. Engineered herbicide tolerance in trees offers arborists and foresters an alternative for weed-management with less cost and more effective control.
The Bt gene encoding an insecticidal protein has been isolated from the bacterium Bacillus thuringiensis, and has already been transferred to a number of agronomic crops. Genetically engineered trees expressing the Bt gene have been obtained in a conifer, Larix decidua, via an Agrobacterium-mediated gene transfer system (44) and in a hardwood tree, Populus, using particle acceleration (58). Bioengineered insect resistant trees should contribute great value to both tree improvement and pest management programs (83). For engineering disease resistant trees, a gene that codes for an antifungal protein, chitinase, which degrades fungal cell walls, is also available (16). Incorporation of this gene may be an important mechanism for controlling plant pathogenic fungi. Resistance against viral infections is possible by incorporating antivirus elements or antiviral genes into trees (87,88). For example, expression of the viral coat protein genes and antisense RNA can protect plants against certain phytopathogenic viruses (5).
Traits for Environmental Stress
Unfavorable soil conditions, water stress, temperature extremes, and atmospheric pollution are important constraints affecting establishment and development of quality trees, particularly in urban environments. Research is underway to understand gene expression and physiological responses to abiotic stresses using conventional and molecular means. For example, genes coding for enzymes such as superoxide dismutase, and osmoregulation genes may be good candidates for conferring tolerance to oxidant (ozone) pollutants and drought/salt stress, respectively. Engineering trees to withstand such environmental stresses will be a long-term goal because the response of plants to stress is a complex process which may involve the expression of more than one gene. However, one should not dismiss the possible contribution of genetic engineering and genome mapping. With recombinant DNA technology, many complex characteristics can be dissected into a number of major determinants and minor determinants (68). Identification and transfer of stress-response traits involving multiple-gene expression has not yet been accomplished; however, success in this area is inevitable. A good example of work with a multi-gene trait is work toward manipulation of wood quality via genetic engineering, discussed next.
Traits for Wood Quality
Modification of lignin structure, composition and content to improve wood quality is being studied in several laboratories. The strategies are generally to isolate and clone several genes coding for a series of enzymes involved in lignin biosynthesis in trees (11,19,91). If this can be accomplished, more desirable wood might be designed via genetic engineering.
Traits for Landscape and Ornamental Purpose
With advances in plant biotechnology, trees can be designed with more desirable ornamental characteristics, such as preferred shapes, sizes, and different colored flowers and leaves. In the past few years, much knowledge about the genetic determination of flower color has been gained at the molecular level. Genes controlling flower color have been identified and cloned, which opens the doors to the development of new ornamental plant varieties (63). Genetic engineering has also made it possible to improve flower longevity, produce more flowers, and modify plant architecture, such as dwarf and upright forms (96). The trait of strong apical dominance in pine has not been introduced into other species by traditional breeding methods. A dominant gene for such a trait may be a transplant candidate gene though it has not yet been isolated.
Genome Analysis and Gene Mapping in Woody Plants
Basic research on the genetics, biochemistry, and physiology of trees has lagged behind that of agricultural crop species. No tree species has been well characterized at the molecular level. However, recent advances in molecular genetics and recombinant DNA technology offer novel tools to explore genetic organization and gene expression in tree species. We will briefly discuss two DNA-based technologies: restriction fragment length polymorphism (RFLP) and random amplified polymorphic DNA (RAPD).
Restriction fragment length polymorphism (RFLP)
This genetic assay depends on the use of a group of enzymes (i.e., restriction endonucleases). These enzymes cleave double-stranded DNA at specific recognition sequences (usually 4-6 base-pair in length). Thus, any base pair or positional changes in the DNA molecule, such as base substitutions, insertions and deletions, will result in a different cleavage pattern, thus variation in restriction fragment lengths. A radiolabeled DNA sequence (called a probe), which hybridizes with a DNA fragment sharing the homologous sequence, is able to detect such variations in length of the DNA fragments. The resultant length variation is referred as a restriction fragment length polymorphism (RFLP). The RFLP assay involves Southern blot hybridization, in which a sample genomic DNA is subjected to enzymatic digestion with restriction endonucleases, size fractionation of the DNA fragments in an agarose gel by electrophoresis, transfer of the DNA from the gel and binding onto a filter or membrane in their original pattern, and hybridization with a radioactively-labeled homologous sequences (i.e., DNA probe), and finally the variation in restriction fragment length (called RFLP markers) is identified on an autoradiogram. Figure 5 shows an example of genetic analysis of a particular trait using RFLP mapping.

An example of genetic analysis of a particular trait in a tree genome using RFLP markers. Two RFLP markers (loci) are identified by a single probe, in which the T locus is a 2.5 kb DNA fragment and the t locus is a 1.5 kb DNA fragment. This diagram represents the hybridization binding pattern of four DNA samples with the probe. The total DNA was isoalated from two parents (P¶ and P2) and their Fļ and F2 progeny, and digested by a restriction enzyme (R).
RFLP, as a tool for genetic analysis at the DNA level, was first used in physical mapping of a temperature-sensitive mutation of adenoviruses in 1974 (36). RFLP mapping on eukaryotic genomes was first described in 1980 by human geneticists (9). In the past few years, RFLP technology has been well developed for DNA-based genetic mapping, genetic diagnostics, molecular taxonomy, and evolutionary studies (93). Today, the RFLP technique has become widely used for the construction of genetic maps of agronomically important species and for the mapping of genetic traits. Linkage mapping using DNA markers is being accomplished in many agronomic crops, such as maize, tomato, wheat, and rice.
Random amplified polymorphic DNA (RAPD)
RAPD technology, developed by Williams et al. (92), produces DNA-based markers like RFLP, but it is based on the selective amplification of DNA segments with a random primer. The RAPD assay relies on the enzymatic amplification of a small amount of target DNA with a single oligonucleotide of an arbitrary DNA sequence (i.e., primer, usually 10-mers) in the presence of a thermostable DNA polymerase. This mixture is subjected to polymerase chain reaction (PCR) under temperature cycling conditions within a machine called a thermal cycler. Then, the PCR products is analyzed by gel electrophoresis. The DNA markers generated from this assay are called RAPD markers.
Currently, these are powerful tools for assaying genetic variation and developing genetic linkage maps. Genetic analyses with RFLP and RAPD markers have quickly been extended to tree species to develop genetic linkage maps. Great effort has being concentrated on mapping two economically important forest species, loblolly pine and poplar. But research activities also include other forest species, ornamental woody plants, and horticultural crops, and are summarized in Table 2. The practical use of these technologies are seen in several areas: using molecular-markerbased selection to improve tree growth response to drought stressed loblolly pine (85), to obtain improved clones having the phenotype of the American chestnut but with the blight resistance trait of the Chinese chestnut while reducing the number of backcross generations (7), to shorten the generation interval in selection for specific gravity improvement (wood quality) in loblolly pine (94), and to identify phytopathogens for disease control in elm species (8).
Woody plants in which molecular markers are being used for genetic mapping and breeding applications.
At Oklahoma State University, we have initiated work studying genetic mechanisms of drought resistance in loblolly and shortleaf pine. Currently our research effort is being focused on locating drought stress and growth quantitative trait loci on a genetic linkage map of loblolly pine using RFLP and RAPD markers (85). Such a linkage map may eventually facilitate marker-aided selection in tree breeding programs and assist in selection for water use efficiency of loblolly pine for genetically improved forest productivity.
RFLP markers have numerous additional advantages over most other types of markers (such as morphological and isozyme). They are developmentally stable, display normal Mendelian inheritance and generally exhibit multiple, codominant alleles (66). Molecular level markers also do not display enviromental effects. RFLP markers are easily detected and relatively easy to map. The major disadvantage of RFLP markers is that they are expensive and time-consuming to generate.
RAPD markers on the other hand, are fairly simply, quickly and inexpensively detected. RAPD markers can be produced with relatively simple protocols and equipment and without the use of 32p, which gives them an advantage over RFLPs as far as application in field labs with limited facilites. They are, however, dominant markers, thus they provide much less information for mapping than do RFLPs. There also are some problems inherent to the PCR technique which can result in “false positives”, which are not useful, and may result in misleading data. For this reason, repeatability should be tested.
Strategies for applications of molecular markers to tree improvement programs
General applications of RFLP and RAPD technologies in plant genetics and crop improvement have already been discussed in several recent reviews (27,53,65,71,84,93). The potential impact of the novel technologies lies in linking classical and molecular genetics. Although RFLP and RAPD markers have been used for a variety of purposes, for tree improvement programs, the special interests and immediate applications may include the following: RFLP and RAPD markers can be used to locate discrete chromosomal loci to a genomic region and to quickly construct saturated genetic maps of individual lines. As genetic linkage (between molecular markers and quantitative traits) maps develop, they can be used to identify, locate and perhaps eventually clone specific traits of interest (such as singlegene abiotic and biotic stress traits), which will be useful for the manipulation of trees by both traditional breeding and genetic engineering. More directly, tree breeders can easily use RFLP and RAPD markers for assessing genetic diversity of natural populations and to study genetic relatedness between individuals (23), choosing parents to design breeding programs, and monitoring gene flow and evaluating segregation in offspring populations (28,12). In addition, DNA marker assays depend upon genome composition rather than protein (isozyme) expression or morphology, which are often influenced by their developmental phase and environment. Therefore, these molecular tools hold the promise to overcome some problems inherent to conventional tree breeding programs.
Summary
Among the various biotechnologies, in vitro techniques and tissue culture are most readily integrated into the improvement of forestry and ornamental woody species. Clonal propagation via tissue cultures is in use today by many commercial growers around the world and is playing an increasing role in forest nurseries and the horticultural industry, yielding relatively high economic return (57). However, the full potential of tissue culture technology has not been realized. Unresolved problems still remain in tree tissue culture research (59), and these problems limit its usefulness as a tool for tree improvement.
Although genetic engineering in forestry and woody ornamental species began only a few years ago, rapid and substantial progress in this research has already been made. Genetically engineered poplar and larch plants expressing agronomically important traits, i.e., herbicide and insect resistance, are under preformance testing. These achievements demonstrate the feasibilty of genetic engineering in woody plant species and have advanced the application of gene transfer and molecular biology to tree species. However, the successful application of genetic engineering in tree improvement requires several steps: isolation of desired genes from trees orotherorganisms, introduction of these genes into the tree genome, and regeneration of whole plants from genetically modified cells or tissues. Gene transfer methods, and particularly, regeneration systems, for many important species are still unavailable, which is an area needing more basic research. At present, few valuable genes from trees have been characterized and isolated. This is probably the most important immediate task for forest molecular biologists in the next 5-10 years. Obviously, genetic engineering of trees will be a long-term research effort and will require continued and increased support from users and consumers. The application of gene transfer techniques will have a major impact on future breeding of woody plants
Forest genetics is still an infant science compared with agricultural genetics. In addition, unlike agronomic crops, forest trees possess large genomes with heterogeneous backgrounds. Therefore, inadequate knowledge of genome structure and regulation of gene expression in trees limits tree breeders and arborists from applying these methods to tree improvement programs. However, rapid advances in molecular genetics and DNA technology contribute to the feasibility of genome mapping in trees. In particular, developing technologies such as RFLP, RFLP-QTL, and RAPD mapping are extending our knowledge of the genetics of trees and are providing the opportunity to understand genes or gene families coding for specific traits. The discovery of RFLP and RAPD markers and use of marker-based selection are very useful tools that can be directly applied to tree improvement programs.
Finally, it should be mentioned that biotechnology and conventional tree breeding are complementary rather than competing techniques. Both are essential components in successful tree improvement programs as illustrated in Figure 6. Therefore, traditional tree breeding methods and modern biotechnology can and should be fully integrated in tree improvement programs (15,73). Such an integration may be more important in the programs for woody plants than herbaceous species because of the former’s often long sexual generations. Genetic engineering in conjunction with traditional methods can provide the means for greater gains and more efficiency in forest tree improvement.

Integration of biotechnology into conventional tree improvement programs.
Footnotes
1 School of Forestry and Wood Products, Michigan Technological University, Houghton, Ml 49931, USA
- © 1993, International Society of Arboriculture. All rights reserved.
Literature Cited
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵
- 81.↵
- 82.↵
- 83.↵
- 84.
- 85.↵
- 86.↵
- 87.↵
- 88.↵
- 89.↵
- 90.↵
- 91.
- 92.↵
- 93.↵
- 94.↵
- 95.
- 96.↵
- 97.↵
In this issue








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.