
Abstract
Field-grown Gleditsia triacanthos seedlings were inoculated each month for two years with the fungus Thyronectria austro-americana. Resulting canker sizes were recorded monthly for one year after inoculation. The season in which inoculations occurred had a significant effect on average canker size. Canker size was largest after summer and fall inoculations, and smallest after winter and spring inoculations. Canker expansion ceased 7 - 10 months after inoculation in the fall, winter, and summer, and 3 - 4 months after spring inoculations. Thus, cankers resulting from spring inoculations were small and only expanded for a few months before host defenses stopped expansion. Cultivars (Skyline, Imperial, Sunburst) and one seedling selection (Thornless) were inoculated at 0, 7, 14, and 21 days after wounding in the spring, summer, and fall of 1986 and 1987. Wounds remained susceptible to infection between 7 and 14 days in all but one season during the experiment. In 1986, spring wounds remained susceptible for 21 days. Thus, the season of inoculation did not greatly affect the duration wounds remained susceptible.
Thyronectria canker of honeylocusts (Gleditsia triacanthos) induced by the fungus Thyronectria austro-americana. causes losses in landscape plantings in many areas of the United States (10,11,12,13,19). Pruning wounds, branch stubs, and openings in the bark, such as branch crotches, are common sites of infection. Many honeylocusts are pruned to improve form and remove problem branches. Disease prevention could be improved if the effect of the season on infection, canker expansion, and the duration wounds remain susceptible to infection were known. This information would allow modification of pruning schedules for more efficient management of the disease.
Previous researchers have found that wounds in several tree species remain susceptible to infection by canker forming fungi for about 7 to 14 days (1,5,9,17,18). However, the time of year, drought, root damage, and degree days after wounding may affect the duration of wound susceptibility (1,14,15). The length of time wounds remain susceptible appears related to the rapidity of suberin and lignin or tannin deposition in cortex cells (3,7). Biggs and Miles (6) found that suberin production was related to resistance in peach (Prunus persica) in May and June, but not in April, July, September, and October.
The season when honeylocusts are most susceptible to infection is unclear. In Illinois, cankers on honeylocust trees inoculated with T. austroamericana in September and November were larger than those on trees inoculated in May or July (16), whereas other researchers (2,8) found more and largercankers resulting from inoculations in June and July than in September inoculations. The duration that wounds on honeylocusts remain susceptible and the season trees are most susceptible to canker inducing fungi in the semi-arid climate of Colorado are unknown. Thus, the objectives for this study were to determine the effect of four seasons (spring, summer, fall, winter) on infection and subsequent canker expansion, and the effect of three seasons (spring, summer, fall) on the duration that wounds remain susceptible to infection by T. austro-americana.
Materials and Methods
Two studies were used to address these objectives. The first was a seasonal (3 months per season) inoculation of seedlings and subsequent monthly monitoring of resulting canker sizes for a year. In the second study, honeylocust cultivars were used to assess seasonal effects on duration of wound susceptibility.
Plant material
Two-year-old honeylocust seedlings (G. triacanthos var. inermis) (podless and thornless) obtained from the Colorado State Forest Service Nursery in Fort Collins were planted at the Department of Plant Pathology and Weed Science Research Nursery in Fort Collins in May 1981. Seedlings averaged 2.4 cm in diameter at 15 cm from the ground and were 1.8 m tall when the seasonal effect on infection study was initiated in September, 1983. For the wound susceptibility study, three honeylocust cultivars (Skyline, Imperial, Sunburst) and one seedling selection (thornless) were planted in 1981 as 2 m high bare-root stock. Cultivars were 4.8 ± 0.9 cm in diameter at 15 cm from the ground and 4.5 ± 0.6 m tall when the study began in 1986.
Fungal material
Three, single-conidial isolates of T. austro-americana, isolated from three different infected honeylocust trees in Denver, CO, were used to inoculate all seedlings in the seasonal infection study. Isolate 83-28, collected from Denver, CO, was used in the wound susceptibility study. Isolates were grown on potato-dextrose agar (PDA) for 10-14 days at 26-28 C, in 8 hrs of light before the inoculation.
Seasonal inoculation
Each month two trees were inoculated with all three isolates, and two trees were used as wounded, noninoculated controls. Inoculations were performed on the 15th of each month for 23 months, starting in September 1983 and ending August 1985. One tree in each inoculated and noninoculated group was partially girdled at 10 cm from the ground by removing a 3 cm wide section of bark from 50% of the circumference. After cleaning the bark with 95% ethanol, three wounds were made by removing the bark with a 12 mm diameter arch punch. The wounds were 10 cm apart starting 10 cm above the girdle or 20 cm above the ground on nongirdled trees. Agar plugs of the three isolates were placed in the three wounds. Sterile agar plugs were used on control wounds. The wounds were wrapped with clear plastic tape, which was removed four weeks later.
Canker size was measured each month for 12 months after inoculation. Total canker size was recorded as the sum of the horizontal and vertical diameters of discolored bark around the wounds. The Manova program in SPSSX (20) was used to analyze differences in average and maximum canker sizes by season, year, isolate, and girdling treatments. The experimental design was a splitsplit plot where year was the block effect, season was the whole plot effect, girdling was the split plot effect and isolate was the split-split plot effect. The month within season by year variance was small relative to the season by year effect, so months were averaged by season. The spring season included March, April, and May. The subsequent seasons each contained three months. Mean separation was by LSD at a P=0.05.
Wound susceptibility
Three trees were inoculated and one tree was used for a wounded, uninoculated control in each of two rows of trees foratotal of six trees per cultivar inoculated. Three cultivars and one seedling selection were used. Each tree was wounded four times on the main stem by removing the bark with a 12 mm arch punch. Wounds were 10 cm apart and at the same height and side on all trees for a particular repetition. The lowest wound was inoculated within 30 minutes of wounding. The remaining wounds were inoculated at 7,14, and 21 days after wounding. Agar plugs containing the fungus were placed in the wound and plastic tape was used to seal the wound. The tape was removed after 2 weeks. The experiment was initiated in the spring (April 15, 1986 and April 28,1987), summer (June 17,1986 and July 10,1987) and fall (October 22,1986 and September 25,1987). The same trees were used in each experiment with the wounds made on a different area of the lower 2.5 m of the stem.
Canker sizes were recorded by measuring two cross diameters of the discolored bark around the wounds. Readings were started at the wounds inoculated on day 0, with the other wounds measured subsequently at 7 day intervals. Readings were taken 30-60 days after inoculation for spring and summer tests and 60-120 days for fall tests. Canker size based on time of inoculation after wounding, cultivar, year, and season was analyzed as a split-split plot design. The two rows of trees were a block effect and the four cultivars crossed with the control and inoculated trees were the whole plot treatments. There were three repeated measure factors including season, year, and time of inoculation. The season (spring, summer, fall) and year were analyzed as split plot factors and time of inoculation after wounding (0, 7, 14, 21 days) was a split-split plot factor. Infections were assumed to have occurred if the area of discolored bark around inoculated wounds was significantly larger than around noninoculated wounds. Mean separation was by LSD at a P=0.05.
Weather data
Weather data from a station one mile from the research plots on the Colorado State University Campus was utilized to assess the possible relationships of canker expansion with monthly average temperature, total monthly precipitation, and deviations of monthly average temperatures from 40 year averages.
Results and Discussion
Seasonal inoculations
The season in which inoculations occurred had a significant effect on canker size. The mean canker size and maximum canker size measured during the 12 months following inoculation were largest after fall and summer inoculations and smallest after winter and spring inoculations (Fig. 1). Infection rarely occurred in the winter (December to February) and when it did, cankers remained small most likely because of the cold weather. In the spring (March to May), cankers formed, but wound closure started after 4 months producing only small cankers (Fig. 1). The large cankers resulting from summer inoculation were unexpected since it was presumed that the tree would inhibit fungal expansion before lower fall temperatures and/or dormancy slows the defense response. In contrast, control wounds of the summer season callused rapidly, with closure within 12 months after wounding. No other seasonal set of wounds did this. Warm summer temperatures may allow rapid growth of the pathogen (17), but it is also reported that warm temperatures promote periderm formation (4). Thus, the reason for the large summer cankers is not known. Larger cankers formed after fall inoculations of year 2 than year 1; this may be attributed to fungal growth during a winter which was 10° F warmer than the 40 yr average. Warmer temperatures could promote fungal growth even while the tree was dormant. The girdling treatment apparently promoted larger cankers to form in year 1 but not in year 2 (Table 1, Fig. 1).

Average canker size on honeylocusts inoculated with T. austro-americana during three monthswithin a season. Half the trees were partially girdled. Means with a different letter indicate a statistically significant difference (P=0.05) among means within girdled treatments and within a year (LSD= 3.0). *Indicates a significant difference (P=0.05) between cankers on girdled and nongirdled trees within season and year (LSD= 2.6).
Number of months between inoculation and maximum canker size on honeylocusts inoculated with Thyronectria austro-americana in different seasons.
Maximum canker size occurred 7-10 months after inoculation in the fall, winter, and summer (Table 1). The shortest time (3-4 months) to maximum canker size occurred when inoculations occurred in the spring. Thus, spring inoculations allowed the shortest time for the fungus to invade tissue before the tree’s defense system restricted canker expansion. Inoculations during the winter of 1984-85 resulted in restricted canker expansion after only 5 months. A warm March, April, and May (5° F above average) may have allowed early host response.
Average canker size varied by isolate, but there were significant isolate, year, season or month and girdling treatment interactions (Fig. 1). These interactions were not explainable biologically, and because the isolates caused similar trends in canker size over the 2 years, canker sizes were averaged across the three isolates.
Wound susceptibility
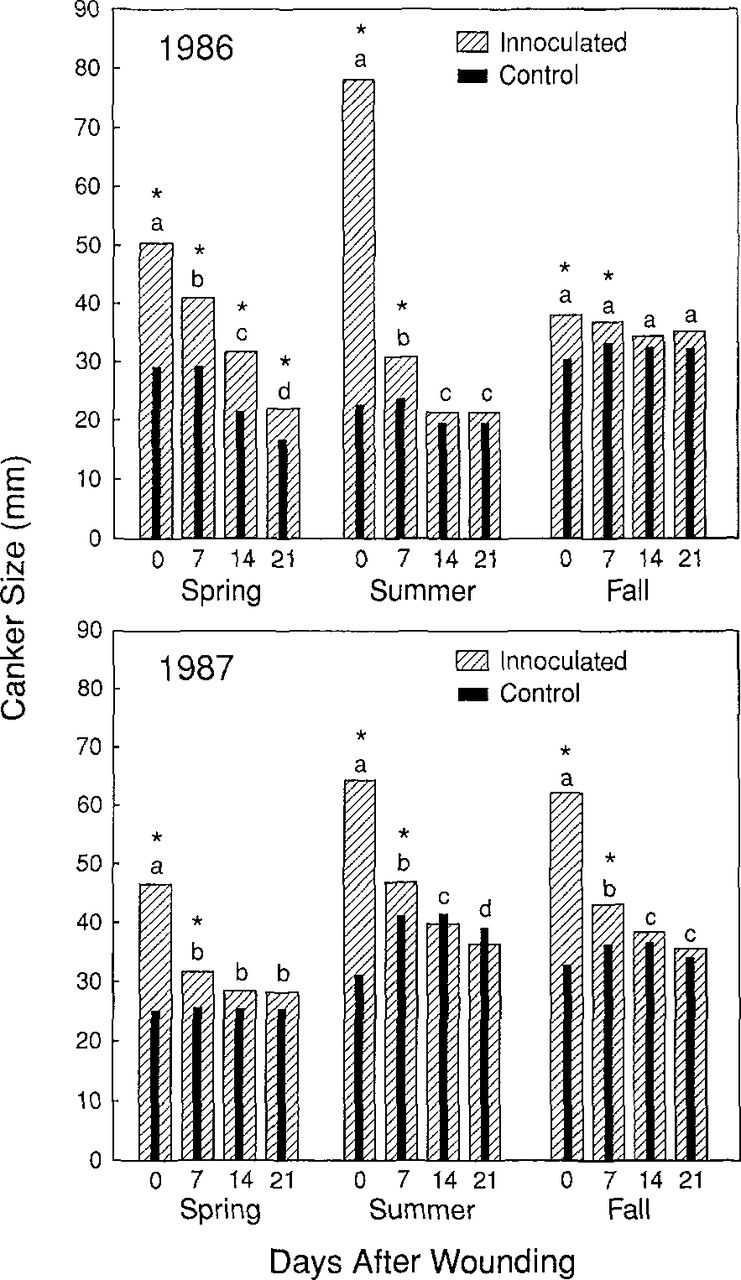
Wounds remained susceptible to infection from day 0 to between 7 and 14 days after wounding during all seasons and years except for the spring of 1986 when infections occurred 21 days after wounding (Fig. 2). Trees in the spring of 1986 may have been stressed from an unusually warm (10° F greater than average for 3 months) and dry winter before inoculation and, thus, may not have been able to resist the fungus. Significantly smaller cankers resulted from wounds inoculated 14 and 21 days after wounding as compared with cankers inoculated on day 0 or 7 for all seasons except fall 1986 and spring 1987.

Average canker size on honeylocusts inoculated with T. austro-americana 0-21 days after wounding in three seasons in 1986 and 1987. Canker means with a different letter indicate a significant difference in means among those inoculated 0, 7, 14, and 21 days after wounding within a season. Based on a LSD of 3.1 (P=0.05). *Indicates a significant difference between controls and inoculated wound size. Based on LSD of 3.6 (P=0.05).
There were significant canker size interactions between years, seasons, control vs inoculated wounds, and time after wounding. These interactions are logical, because inoculations occurred in different months within a season and there was variation in the length of time between inoculation and determination of canker sizes. Thus, data were analyzed as six separate experiments instead of examining data over time. Cultivars did not affect canker size in these tests although cultivars vary in resistance (14,16). Based on these relatively large time intervals of 0, 7, and 21 days, season apparently did not affect the duration that wounds remained susceptible to infection. Probably, this was because all inoculations were completed during months of warm daytime temperatures. Thus, even fall wounds dried out and became poor infection courts because it was likely warm enough for host defense reactions to occur in fall and spring. The story is not altogether so simple because, under some conditions, wounds apparently stay susceptible for 21 days. Further studies are needed before we can determine the effect of various weather induced stresses on honeylocust’s resistance to infection.
Means of reducing losses to Thyronectria canker include appropriate timing of pruning, reducing stresses, and increasing host resistance. Avoidance of pruning or wounding honeylocusts in late summer, fall, and winter would reduce the duration in which cankers could expand before host defenses can contain the fungus. Under normal growing and environmental conditions, wounds become resistant to infection between 7 and 14 days after wounding. However, spring pruning should be avoided if the activity causes more injuries than at other seasons. To reduce infections, pruning during any season should be done when there are at least seven days of low humidity and no precipitation. Treatment of wounds when they are susceptible for 1-2 weeks with fungicides is not effective at legal rates (Jacobi, W. R. Unpublished data, 2). Trees under various environmental stresses, such as severe fluctuations in soil moisture, however, may be restricted in defending against infection, and wounds may remain susceptible to infection for longer periods. Other means of reducing losses could include the use of resistant cultivars (14,16). Continuing studies at Colorado State University are addressing the issue of fluctuating soil moisture as a predisposing stress.
Acknowledgments
I thank J. R. zumBrunnen of the Statistical Laboratory, Colorado State University for statistical advice and J. P. Hill, H. F. Schwartz, G. A. McIntyre, and E. G. Ruppel for reviewing this manuscript. Funding forthis research was provided by Colorado Agricultural Expt. Sta. and International Society of Arboriculture. Donation of plant material by Baily Nurseries is appreciated.
Footnotes
↵1. A publication of the Colorado Agricultural Experiment Station, Journal Number 153451.
- © 1992, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.