
Abstract
Root-zone temperatures in city landscapes are supraoptimal for the growth of many temperate tree species, and selection of heat-resistant genotypes will aid in the development of successful urban forests. The purpose of this study was to compare the growth of Gleditsia triacanthosvar. inermis (honeylocust), Maackia amurensis (Amur maackia), and Sophorajaponica (pagoda tree) athigh root-zone temperature. Nutrient solution in which seedlings were grown was held at 35°C, for 0, 6,12, and 24 hr/day for 42 days. For honeylocust and Amur maackia, dry mass was similar for plants at 35°C 0 and 6 hr/day but was reduced among plants at 35°C for 12 and 24 hr/day. In contrast, mass of pagoda tree was decreased 55% by 35°C root-zone temperature for 6 hr/day, but mass increased as exposure to 35°C increased beyond 6 hr/day. Young lamina of honeylocust and Amur maackia at 35°C for 12 and 24 hr/day were chlorotic, and a high-temperature-induced reduction in the concentration of iron in leaves of all species was confirmed. These results indicate that growth of pagoda tree seedlings is more sensitive to cyclic episodes of high rootzone temperatures than to constant heat. The influence of high root-zone temperature on growth of Amur maackia was similar to that of honeylocust, which in previous studies was more heat resistant than other trees used in temperate urban forests.
High root-zone temperature affects the growth of woody plants both during production and after installation in the landscape. Temperatures exceeding 4O°C for up to 6 hr/day can occur in soils of plants produced in containers (13). Root-zone temperatures of urban street trees can exceed 3O°C, and the average soil temperature near street trees was 7°C higher than the soil temperature in an adjacent temperate forest (4). Because many species of woody plants used in urban landscapes evolved in environments where soil temperatures are moderate, selection of genotypes resistant to high root temperature will aid in the development of successful urban forests.
Resistance to high root-zone temperature has been assessed in only a few species of temperate landscape trees. Only in seedlings of honeylocust (Gleditsia triacanthosvar. inermis) has the growth at a root-zone temperature below 3O°C been sustained with continuous exposure to temperatures as high as 34°C (7). Honeylocust is one of the most frequently planted trees in stressful urban landscapes, but several other members of the Leguminosae family appear to have merit for increased use in cities. Among these species are Amur maackia (Maackia amurensis) and pagoda tree (Sophora japonica), but no information is available on the responses of these species to high temperatures and other potential constraints on growth in urban tree microclimates. The purpose of this study was to compare the growth of seedlings of honeylocust, Amur maackia and pagoda tree exposed to high root-zone temperature.
Materials and Methods
Half-sib seeds of honeylocust, Amur maackia, and pagoda tree were scarified in 18MH2SO4 for 45, 60, and 60 min, respectively, and germinated between sheets of paper moistened with tap water at 25°C in the dark for 7,10, and 10 days, respectively. One uniform seedling of each species was placed in each of 36 1.2-liter stainless steel beakers filled with 10% Hoagland solution #1 (12) modified to contain 50μM phosphorus (P). Iron (Fe) was supplied as Fe-EDTA, and pH was increased to 7.0 with KOH. These modifications improved the growth of Amur maackia with no effect on honeylocust and pagoda tree during preliminary experiments. Solutions were aerated continuously, replenished once daily with deionized water, and replaced every 7 days throughout the experiment.
Each beaker was suspended in an individual polyvinyl chloride jacket allowing temperature-regulated water to be circulated around the outer steel walls of the beakers for varying amounts of time each day. This system for controlling rootzone temperature, based on that described by Graves and Dana (5), was housed in a plant growth chamber in which air temperature was held at 22 (±1.5)°C, cool-white fluorescent lamps upped 16-hr photoperiods of 280(+30) μmol/m2 / s (400-700 nm), with no relative humidity control.
After 3 days during which root-zone temperature in all beakers was held at 23(±2)°C, seedlings were treated by raising solution temperature to 35(±0.5)°C for either 0, 6, 12, or 24 hr/day. Heat treatments for the 6 and 12 hr/day regimes were applied during hours 8 through 13 and 5 through 16 of the photoperiod, respectively. About 4 hr were required for heated solutions to return to 23°C after daily episodes of high temperature.
Plants were harvested after 42 days of rootzone temperature treatment. Dry mass of leaves, all other shoot parts, and root systems was measured after drying tissues in a forced-air oven at 65°C for 2 days and allowing samples to cool to room temperature for 1 hr. Iron (Fe) content of lamina was determined using an Inductively Coupled Plasma-Atom Emission Spectrophotometer. This experiment was performed twice, and data were combined for analysis. The significance of linear and quadratic effects in the relationship of dependent variables to exposure to 35°C root-zone temperature was determined by regression using the Statistical Analysis System.
Results and Discussion
The influence of increasing exposure to 35°C root-zone temperature on total dry mass was similar in seedlings of honeylocust and Amur maackia (Fig. 1 A). High temperature for up to 12 hr/day had little effect on dry mass in both species, but, compared to seedlings not treated with heat, the mass of seedlings at 35°C for 24 hr/day was reduced 56 and 54% in honeylocust and Amur maackia, respectively. In contrast, 35°C root-zone temperature for 6 hr/day decreased the dry mass of pagoda tree seedlings by 55%, and the mean dry mass of pagoda tree seedlings treated 24 hr/day with high root-zone temperature was 29% greater than the mass of those at 35°C for 6 hr/day (Fig. 1A). High root-zone temperature increased the ratio of lamina (leaves without the pediole) to root dry weight in honeylocust and Amur maackia, but this ratio tended to decrease with increasing exposure to high temperature for seedlings of pagoda tree (Fig. 1B).
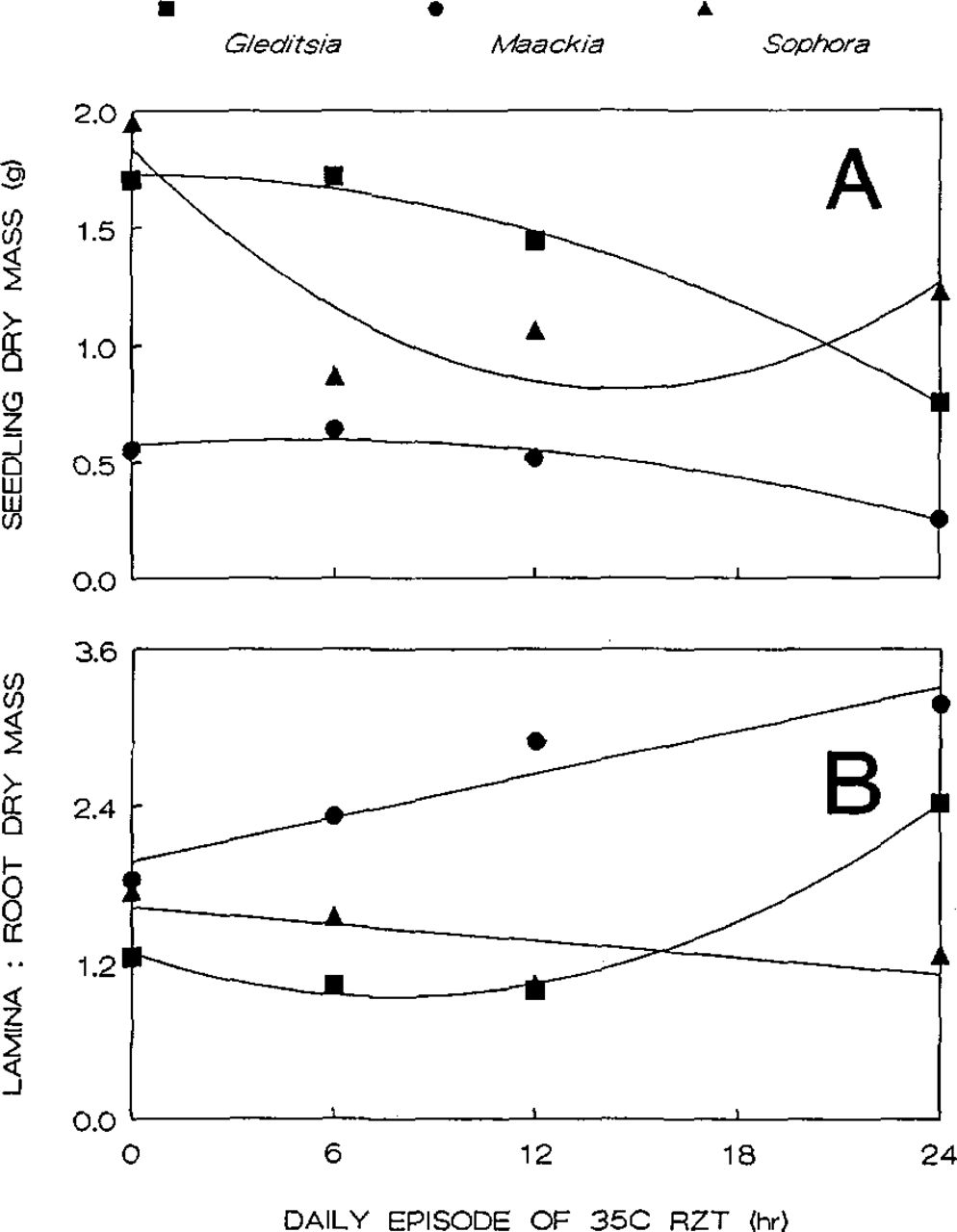
Dry mass (A) and the ratio of lamina to root dry mass (B) of honeylocust, Amur maackia, and pagoda tree seedlings grown in solution and exposed to 35°C root-zone temperature for 0, 6,12, and 24 hr/day for 42 days. Each point represents the mean of 18 replicates. Regression functions for A: Y = 1.73+0.0003(X) - 0.002(X2) [R2=0.993)] (Gleditsia); Y = 0.57 + 0.01(X) - 0.001(X2)[R2 = 0.961] (Maackia); Y = 1.84 - 0.14(X) + 0.005(X2)[R2 = 0.777] (Sophorá). Regression functions for B: Y = 1.28 - 0.09(X) + 0.006(X2)[R2 = 0.994] (Gleditsia); Y = 1.97 + 0.06(X)[R2= 0.910] (Maackia); Y ≡ 1.63 - 0.02(X)[R2 = 0.482](Sophora).
Root-zone temperature effects on growth and dry matter partitioning of honeylocust are consistent with those reported by Graves and Wilkins for solution-cultured seedlings grown from seeds harvested from the same tree during the same year as the seeds used in this study (6). It is interesting, however, to compare these results to those of a prior study in which honeylocust plants were grown with either 24 or 34°C root-zone temperature for 24 hr/day (7). In that study, the dry mass of seedlings at the two temperatures was not different. Several factors may explain these results. The 1°C difference in the root-zone temperature used in these experiments may have been sufficient to cause a marked difference in growth. In general, plant growth increases gradually with increasing temperature until some threshold is reached, above which growth decreases precipitously (10). A root-zone temperature of 34 to 35°C may represent such a threshold for honeylocust. The seedlings used in the earliest study (7) also may have differed genetically from the seedlings used here and in the experiments by Graves and Wilkins (6), causing the heat tolerance of seedlings from the two seed sources to differ. Finally, differences between the growth media used in these studies may have affected the capacity of the seedlings to avoid or tolerate heat. Additional research is needed to address these possibilities.
Seedlings of honeylocust and Amur maackia, for which 35°C root-zone temperature for 12 hr/day caused reductions in total dry mass of 15 and 7%, respectively, appear to be more resistant to root-zone heat than several other temperate tree species. For example, Martin etal. (14) found that the shoot dry mass of Ulmus parvifolia ‘Drake’ plants with 35°C root-zone temperature for 6 hr/day was 39% less than the shoot dry mass of plants at 28°C for 6 hr/day. Graves et al. (9) reported that the shoot dry mass of Acer rubrum grown with continuous 36°C root-zone temperature was 57% less than that of plants with 24°C root zones. Shoots of Ailanthus altissima seedlings grown with a root-zone temperature of 36°C consisted of 47% less dry matter than shoots of seedlings grown with 24°C root zones (8). That the dry mass of seedlings of pagoda tree with 35°C root-zone temperature for 6 hr/day in this study was 22% less than that of those treated for 24 hr/day indicates that seedlings of this species are more sensitive to cyclic exposures to rootzone heat than to continuously high temperatures. The mechanisms underlying this response, which was observed in both runs of this experiment, merit further attention.
Although daily episodes of 35°C root-zone temperature caused marked decreases in the mass of pagoda tree seedlings, the lamina of these plants were green throughout treatment in all temperature regimes. This was in contrast to lamina of honeylocust and Amur maackia plants exposed to root-zone heat. Lamina near the stem apices of these plants appeared chlorotic after about 3 weeks of treatment, most notably among plants exposed to 35°C root-zone temperature 12 and 24 hr/day. The localization of chlorosis in young tissue suggested that the seedlings were deficient in Fe. Subsequent analysis of Fe content of lamina supported this hypothesis (Fig. 2). Fe content of pagoda tree seedlings also was reduced by 35°C root-zone temperature. However, lamina of pagoda tree seedlings contained more Fe than lamina of the other two species, and the appearance of pagoda tree foliage indicated that the amount of Fe in young leaves was sufficient to sustain the synthesis of chlorophyll.

Iron in lamina of honeylocust, Amur maackia, and pagoda tree seedlings exposed to 35°C root-zone temperature for 0,6,12, and 24 hr/day for 42 days. Each point Is the mean of two replicates of one to four subsamples that consisted of all lamina from all plants combined within each species-temperature treatment combination. Regression functions: Y=112.9 - 9.3(X)+0.2(X2)[R≡=0.984)] (Gleditsia); Y = 79.0 - 6.2(X) + 0.2(X≡)[R2 = 0.999] (Maackia);,Y = 189.1 - 18.9(X) + 0.6(X2)[R2= 0.942] (Sophora).
There is little information in the literature on the influence of high root temperature on Fe uptake. In this study, Fe was supplied in nutrientsolution as Fe-EDTA, in which Fe in the ferric form (Fe3+) is bound to the chelate EDTA. Roots take up ferrous Fe (Fe2+) after Fe3+ in the chelate complex is reduced and released from the chelate (2). High temperature may interfere with the reduction of Fe3+ or the transport or metabolism of Fe2+. Alternatively, EDTA may have been rendered ineffective by heat or changes in solution pH during this study. This possibility seems unlikely because we have obtained similar results for heat-stressed honeylocust seedlings provided Fe-EDDHA (data not presented). The stability of EDDHA is much less sensitive to pH that EDTA (11), and there is no evidence that the properties of either chelator are affected by the temperatures used in this study.
Summary
These results illustrate effects of high rootzone temperature on seedlings of honeylocust, Amur maackia, and pagoda tree. Root-zone temperature affected the development and partitioning of dry mass similarly in honeylocust and Amur maackia. Previous research indicates that honeylocust is more resistant to high root-zone temperature than many other trees used in urban landscapes where high soil temperatures prevail. Assuming temperature responses in seedlings are comparable to those in mature plants, the similarity between Amur maackia and honeylocust in the capacity to grow at high root-zone temperature indicates that Amur maackia merits increased use in city landscapes with high soil temperature. Its form and ultimate size indicate that Amur maackia could be used as a street tree (3). This study suggests that trees of this species would be comparably resistant to the high rootzone temperatures at street tree planting sites (4), and the capacity for this species to fix atmospheric nitrogen recently was confirmed (1). However, further investigations should determine the extent to which high soil temperatures cause nutritional imbalances in Amur maackia and other trees in the landscape. In contrast to honeylocust and Amur maackia, growth of pagoda tree seedlings was reduced more by daily cycles of 35°C rootzone temperature than by constant heat treatment. This species may be particularly sensitive to extreme temperature fluctuations in containers (13) and raised planters. Further studies to characterize the relationship between time of exposure to root-zone heat and pagoda tree growth, as well as the mechanism responsible for the reduced Fe content of lamina of all species at high temperature are warranted.
Acknowledgments
The technical assistance of Wilhelmina van de Poll, Nhut Nguyen, and Carrie Green is acknowledged gratefully.
Footnotes
1 Scientific article A6195, Contribution 8364 from the Maryland Agr. Expt. Sta. This research was supported by the Maryland Agricultural Experiment Station’s Competitive Grants Program.
- © 1991, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.