
Abstract
A water soluble dye, methyl violet, was injected into the stems beneath branches of trees representing eight deciduous species at various times between April and October, 1989. The stems and branches were then excised, the bark removed, and the pattern of dye translocation traced. Dye injected directly beneath a branch moved distally straight into the branch. Dye injected beneath, but not directly beneath, a branch moved into the sides or top of the branch or into the stem. When dye was injected beyond the branch shoulder, it moved around the branch but remained in the stem. Patterns of dye movement did not vary among species or with time of year. The results do not support the hypothesis that trunk collars form over branch collars each year at the stem-branch juncture.
The raised or swollen area at the base of a branch in woody angiosperms has been referred to as the shoulder ring (1), branch collar (6), or trunk collar (6). The hydraulic architecture at the branch junction is not clearly understood or described. In a classic diagram, Eames and McDaniels (1) show a direct connection of xylem tissue on the top of the branch to stem xylem directly above the branch. Shigo (8) established that there was little or no direct structural or conductive tissue in the branch crotch. He illustrates the abrupt turning into the branch of stem xylem from below the branch (6). Lev-Yadum and Aloni (2) describe circular nonfunctioning xylem vessels in the branch crotches of oak and pine. The “ball and socket” arrangement in trees is readily observed when branches break under snow or ice loads.
The anatomy of the branch base has become a basis for justifications on where branch pruning cuts should be made. Neely (3,4) has shown that cuts through the shoulder close in a shorter time, and he considers them aesthetically more pleasing than cuts made further out on the branch. In target pruning, Shigo (5,7) recommends that the final cut be made outside the collar. He states that severing of trunk tissue can result in discoloration or decay of the stem. Shigo (6,8) has illustrated an alternative view of branch attachment. In his hypothesis, branch xylem that develops early in the season forms a branch collar, and trunk xylem that develops later in the season forms a trunk collar. The objective of the experiment described here was to observe the water transport system at stem-branch junctures in eight species of woody angiosperms by injecting a translocating dye into tree stems.
Materials an Methods
The experimental trees were growing in the Illinois Natural History Survey arboretum, 1 mile south of Urbana. The trees were 6 to 20 years old, of uniform age within species, and of seedling origin. Eight species were tested. Sugar maple (Acer saccharum), black walnut (Juglans nigra), tulip tree (Liriodendron tulipifera ), Sargent crab (Malussargentii), and basswood (Tiliaamericana) are diffuse porous; green ash (Fra×inus Pennsylvania), honey locust (Gleditsia triacanthos var. inermis), and English elm (Ulmus procera) are ring porous.
Treatments were made at various times from before leaf emergence until first frost: April 25, May 9 and 25, June 20, July 17, August 31, September 15, and October 5, 1989. On each date, two vigorously growing, primary stems were selected on a tree from each of the eight species under investigation. Three holes, each 1 mm in diameter and 5 mm deep, were drilled into the xylem of the stem of each tree 2 to 3 cm below an attached branch. One hole was centered proximal to the branch base (X in Fig. 1) and the other two at sites equidistant from the first and approximately proximal to the flaring branch shoulder (D1 and D2 in Fig. 1). Mean stem and branch diameters are given in Table 1.
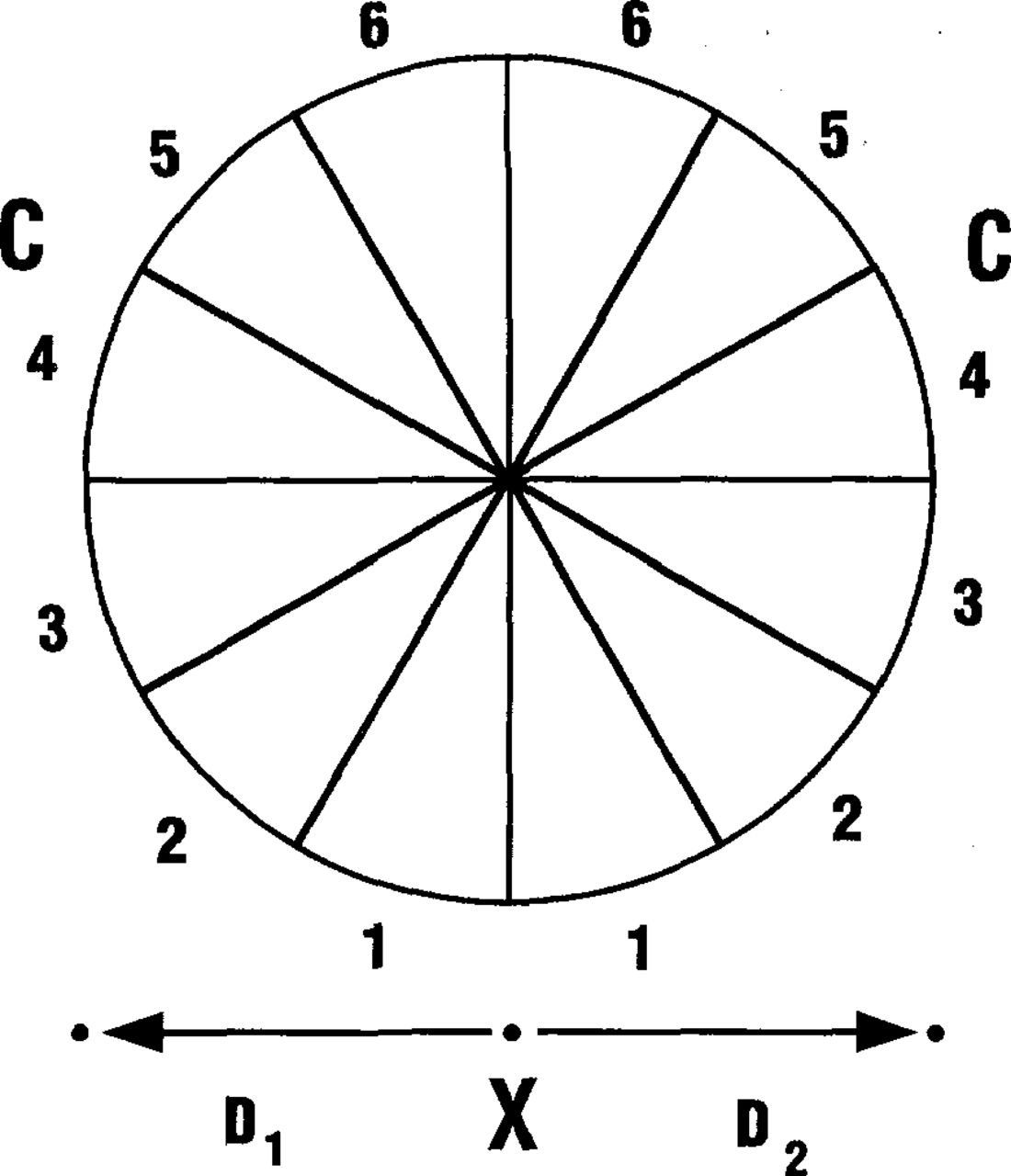
Diagrammatic view of a transverse section through a branch. Dye was placed into three holes in the stem below the branch (X, D1 and D2). When the dye moved into the branch, it discolored one or more sectors. The number (1 through 6) of the discolored branch sector (S) was recorded for each injection
Branch and stem diameters and distal distribution of dye from the injection sites beneath or somewhat outside the shoulder of the branch
Methyl violet (5 × 10-3 ml) was inserted into the drilled holes with a hypodermic syringe. Five to 7 days later the stem and branch were excised from the tree and taken to the laboratory. The bark was removed from the wood, and the dye movement patterns were observed and recorded. The transverse section through the branch (Fig. 1), was divided into 12 equal sectors. The location and width of the xylem pathway as determined by dye movement into the branch from the D1 and D2injection sites were recorded as 1 through 6 using the branch sector arcs. Arc numbers increased from the ventral to the dorsal surface of the branch. Dye movement only in the stem was recorded as a 7.
A supplemental study was conducted in 1990 in which dye was injected in the same fashion into stems beneath dead or severely declining branches. Two branches on a tree from each of the eight species were treated in August. For photographic purposes, an additional stem of basswood with four vigorous branches was treated with a series of injections in August. The basswood stem was 8.5 cm in diameter and the injection sites were approximately 8 mm apart in a row beneath a branch. After 7 days the stem was excised, the bark removed, and the pathways of dye movement observed. To improve the quality of black and white photographs, the dye patterns were highlighted with ink.
Results
The diameter of the branch shoulder always exceeded the diameter of the branch itself, and the diameter of the stem was greater than that of the attached branch. Mean diameters of the stems and branches are given in Table 1, as are the ratios of shoulder width to branch diameter. Branch shoulders were approximately 40 to 50 percent wider than their respective branch.
Dye moved distally from the insertion site and was easily observed after each injection. Air embolism had no apparent disruptive effect on dye uptake. The three major pathways of dye movement were into the branch, into the stem, and into both the branch and stem. Dye was present almost exclusively in the vascular cambium or in the xylem produced during the current season. When present, discoloration occurred throughout the outermost growth increment. Lateral translocation of the dye was limited. Pathways of dye movement were similar throughout the year. Dye from the center hole (X in Fig. 1) moved into the branch in every species at every date.
The pathway of dye from the lateral injection sites (D1 and D2 in Fig. 1) was into the branch, into the stem, or into the branch and the stem. Data onpathways of dye distribution are presented in two ways in Table 1. Combined data from 256 lateral injections (all dates, all species) indicate that dye from approximately 45 percent of the injection sites remained in the stem and dye from the remaining 55 percent moved into the branch or into the branch and stem. Data from 179 of those injection sites located beneath the branch shoulder (lateral distance of 0.5 to 1.5 times the branch radius) indicate that dye from 69 percent moved into the branch. Dye from 18 injection sites measured to be not beneath the branch shoulder (D>1.5 times the branch radius) also was transported into the branch.
The pattern of distribution of dye did not vary appreciably with species, time of year, or type of porosity. The number of lateral injection sites to the left (D1) or right (D2) that were beneath the branch shoulder (determined as 0.5 to 1.5 times the branch radius) for each tree species and each injection date are given in Table 2. The lateral distance of the injection site from directly beneath the branch (× to D1 or to D2) was directly correlated with the position of the dye in the branch (1 to 6 in Fig. 1) or in the stem (position 7). The correlation coefficients of these distances for each species and each injection date with the site of ultimate dye deposition in the branch also are given in Table 2. The correlation coefficients usually ranged from .50 to .70, not highly significant but always positive. Basswood with the largest stems and the largest branches gave the best correlation. Dye patterns from injections in late summer or early fall did not differ from those earlier in the year and the correlation coefficients were similar. Dye injected into the stem beneath the branch shoulder consistently entered the branch (Fig. 2). Dye injected into the stem just beyond the branch shoulder moved around the branch shoulder, laterally just above the branch, and then distally straight up the stem. Dye injected into the stem well beyond the branch shoulder moved distally straight up the stem. Exceptions to the above pattern occurred in branches that were in a severe state of decline or dead. In such branches, all of the dye injected directly beneath the branch shoulder moved laterally around the branch and then distally straight up the stem (Fig. 3).
Number of injection sites and correlation coefficients of the left and right injection site distances (Fig. 1) to the distal distribution of dye in the circumference of the branch.

Effect of injection site on pattern of dye movement in vigorously growing Tilia americana. A. Dye injected into the stem directly below the branch moved distally straight into the branch. B and C. Dye injected into the stem somewhat laterally from directly below the branch moved distally into the branch but to the sides of the branch. With greater lateral injection site distances, more of the dye moved to the side and top of the branch. B and D. When the injection site was not beneath the branch shoulder, the dye moved distally only in stem xylem. Curving dye distribution patterns illustrate that stem tissues directly above the branch crotch were in the stem sap stream. E and F. When the lateral distance of the injection site was intermediate, dye was distributed into the connective tissue in the stem crotch and into both the stem and the branch. (Dye patterns were highlighted with ink for photographic purposes.)

Effect of injection site on pattern of dye movement around dead branches. A. Acer saccharum. B. Liriodendron tulipifera. (Dye patterns highlighted with ink for photographic purposes.)
Discussion
In tests in which a dye was used to locate the most recently formed xylem produced by the vascular cambium in deciduous trees, no evidence of change in the pattern of xylem formation was found in any of the eight species throughout the growing season. The pattern changed only when the branch died or was in a state of severe decline. The evidence, therefore, does not support Shigo’s (1985) hypothesis that trunk collars form each year over branch collars at the stem-branch juncture. If Shigo were correct, the dye injected late in the season would remain in the stem.
Histological justification to support the annual branch and trunk collar hypothesis may be difficult to find. Increase in stem diameter in woody plants results from meristematic activity by the vascular cambium to produce wood and bark. The cambium forms a continuous thin sheath over the stem, branches, and roots except for the growing tips of stems, branches, and roots (Eames and McDaniels, 1947). It functions from the time of its formation until the plant or plant part dies. The meristematic cells of the cambium either are iso-diametric and give rise to vascular rays, or are elongated, fusiform cells that divide to form tracheids, vessels, sieve tubes, and other vertically elongated elements. The ratio of length to width of the elongated cells in deciduous trees in tangential section may be 5 to 1, 25 to 1, or greater (Eames and McDaniels, 1947). Division of cambial cells occurs primarily in one plane (periclinal). An annual conversion of xylem tissue from that extending into the branch to that of xylem tissues extending up the stem would consequently require major cellular contortions by the vascular cambium.
Branches on trees, other than those from adventitious or secondary buds, originate from primary tissues near the twig tip. A branch, therefore, is as old as the stem to which it is joined. As successive annual rings of xylem are laid down by the cambium, the tissues within the cylinder of both the stem and its branch are buried more and more deeply (Eames and McDaniels, 1947).
The stem is larger in diameter than its branch, consequently, the annual rings of xylem in the branch are smaller than those in the stem. The xylem receives elaborated food materials from the adjacent phloem, primarily through vascular rays. The base of the stem benefits from its location by using food materials from both the branches and the stem and thus is larger than the branch itself. This growth pattern creates the branch shoulder.
The diagram of branch burial by Eames and McDaniels (1947) is incorrect because the stem xylem tissue distal to the center of a branch is not connected directly to the dorsal tissue of the branch (Shigo, 1985). The two-dimensional diagram of the stem:branch interface should be similar to that of a leaf trace in a woody perennial plant (Eames and McDaniels, 1947, Fig. 82, p. 181). The connective tissue in the branch and stem crotch receives nutrients and water from the proximal xylem tissues at the margin of the branch base.
Collars on branches form as callus tissue of the stem attempts to overgrow dead or declining tissue of the branch. As a result, branch collars are present only around weakened or dead branches and are distinct from branch shoulders. Branch shoulders, on the other hand, are present at the base of every vigorously growing branch.
- © 1991, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.