
Abstract
The efficacy and temporal pattern of gypsy moth mortality resulting from implantation and injection of pin oaks with synthetic organic (acephate) and microbial (gypsy moth nucleopolyhedrosis virus = NPV, and Bacillus thuringiensis Berliner = B.t.) insecticides was investigated. All mortality occurred in the larval stage. Implantation of acephate produced significant increases in cumulative mortality. Neither microbial treatment showed a significant increase in mortality over controls. In all acephate treated trees, and in one of three B.t. treated trees, there was an exponential sigmoid pattern of increase in cumulative mortality over time. There was considerable variation among trees. In the acephate treatment, this was expressed as variation in the lag-time of a sigmoid mortality pattern. In the B.t. treatment, larvae on only one of three trees showed a mortality response. The successful use of implantation of acephate in this and other recent papers suggest that this technology will undoubtedly have a place in gypsy moth integrated pest management programs in urban environments. Further work to determine the reasons for the lack of success with the microbials is warranted.
In North America, the gypsy moth, Lymantria dispar has defoliated 0.2 million hectares annually since 1969, ca. 0.4 million hectares in 7 of 11 years since 1969, and over 4.9 million hectares in 1982 (1). Since its introduction into Medford, Mass, in 1869, established populations have spread southward into Virginia and westward into Michigan. Large populations have also been recently subjected to massive eradication efforts on the western coast of the US. While most defoliation can be expected to occur on relatively sparsely inhabited forest land, urban and suburban forests and individual shade trees are also defoliated. Amenity trees are particularly important because they substantially increase real estate values and affect the climate and aesthetics of residential communities (2, 3, 4).
Suppression of gypsy moth populations in recent management programs has included aerial or ground applications of synthetic or microbial insecticides, intensive trapping, augmentation of parasitoids, and mating disruption (5, 6, 8). Several contact poisons or contact and stomach poisons are registered for use against the gypsy moth. One compound, acephate, has been shown efficacious for gypsy moth control using tree implant or injection techniques (10). Implantation or injection application may be useful for individual high-value trees in residential communities due to the minimization of insecticidal drift and ease of use by homeowners. Currently, implantations of acephate is registered for use by homeowners (general use category), and formulations directed against gypsy moth applied with injection technologies are in the restricted use category.
Two microbial insecticides, Bacillus thuringlensis Berliner var. kurstaki (B.t.), and a nucleopolyhedrosis virus (NPV), are also used via foliar spray application against gypsy moths. These compounds are bacterial and viral agents, respectively, and require ingestion and biological activity or activation in the insect to be effective. Current application technology for these compounds requires ingestion of material on leaves. If systemic application techniques were possible the probability of ingestion of a lethal dose could be greatly enhanced due to the presence of the microbial in leaves and the reduced probability of phytodegradation or washoff from rain. Microbial insecticides may be preferred over synthetic organics in areas of high environmental sensitivity, such as residential communities.
A common concern with the use of microbials is the time required for mortality. In contrast to a mode of action that depends on interruption of physiological processes following contact, as in many synthetic organic compounds, microbials require biological activity in insect tissue following ingestion. Although feeding may cease in infected larvae prior to mortality, the relatively quick knockdown of synthetic insecticides makes them more appealing to some professional arborists.
This study was designed to investigate systemic application of microbial insecticides directed against the gypsy moth. In addition, the time required for mortality when using systemic application was measured for both microbials and acephate, a synthetic organic insecticide.
Methods and Materials
A replicated complete block experiment using three blocks and three insecticide treatments plus a control was conducted. The insecticide treatments were acephate, B.t., and NPV. Untreated trees served as controls. Treatments were applied to pin oak (Quercus palustris) trees growing on the edge of an approximately 15-year-old stand. Trees had a mean (±SE) dbh of 14.3 ±2.0 cm (n = 12). The pH of the sap was measured on April 14, 1988, by using pliers to express sap from peeled twigs onto a series of pH papers. Treatments were timed to coincide with early leaf expansion and early gypsy moth larval development. This timing was presumed to optimize uptake in trees and to expose the most vulnerable gypsy moth life-stages, the early larval instars, to the insecticide. Treatments were applied on May 13 and 14, when leaves at the top of the trees were ca. 40% expanded and leaves at the bottom of the tree were ca. 20% expanded. Larvae were ca. 13 days post eclosion (egg hatch) at 24°C at this time (see below).
The acephate treatment was applied as implants of a powdered formulation (97%ai) where each implant contained 0.902 g of formulation (0.875 g ai). The insecticide was contained in gelatin capsules within plastic cartridges (ACECAPSR) (Creative Sales, Fremont, NE). Implantation materials and procedures were those used by Webb et al. (10) except that they timed the implantation at or soon after budbreak. A 4 cm deep by 0.95 cm diameter hole was drilled into the bole at the rate of one hole per 10 cm of trunk diameter (4 to 5 implants per tree in this experiment) in a spiral pattern starting near the base of the tree. An ACECAP was inserted and tapped to the bottom of each hole with a metal rod and hammer. The cartridge was not broken in this process.
Microbials were stored frozen and suspensions made on the day of injection. Suspensions were made in pH 5.6 deionized water and protected from sunlight. Both microbial insecticide treatments (B.t. and NPV) were applied by injection into flare roots (7) through six ports per tree using ca. 10 to 12 psi pressure to try to introduce a four liter suspension into the tree. We initiated all injections during the mid-morning to late afternoon hours, and continued the injection process until the suspension was completely taken up or no longer being taken up. The injection system continuously recycled the suspension to inhibit sedimentation.
For the B.t. treatment, the washed spores and delta-endotoxin protein crystals from 3.75 grams of B.t. var. kurstaki powder (Abbott Laboratories, Chicago, IL) were suspended in four liters of deionized water for each tree. Injection of the first B.t. replicate resulted in a great deal of foaming and we were unable to put the entire treatment into the tree (see results below). In an effort to reduce this problem, a rotary knife blender was used to homogenize the suspension in the latter two replicates.
The NPV insecticide was a formulation of nuclear polyhedrosis inclusion bodies (PIBs) cleaned so as to include only PIBs in the 3 to 5 micron diameter range. Trees were injected with a four liter suspension at a rate of 1.5 × 109 PIBs per liter. All suspensions were homogenized for three 30-second time spans in a rotary knife blender.
Efficacy and the temporal pattern of mortality were measured using a bioassay against gypsy moth larvae. Gypsy moth eggs were collected from a building population at Gimlet Ridge, Shenandoah National Park, Va., surface sterilized in 10% formalin for 1 hour, and rinsed thoroughly in tap water. Hatched larvae were transferred to 3.9 liter waxed paper cartons with screened tops at a density of 20 larvae per carton. Five cartons were made for each tree (15 cartons per treatment). Cartons were held in an environmental chamber, at 14:10 L:D, 24°C. An open pan of water was placed in the chamber to maintain humidity. During early development larvae were fed pin oak or northern red oak, Quercus rubra, depending upon availability of foliage.
Starting 1 to 2 days post tree injection and continuing until pupation, larvae were provided with foliage only from the experimental trees. New foliage was provided as bouquets in vials of water at one to two day intervals. This bioassay method allowed larvae to complete development or die on treated or control foliage.
Cumulative mortality of larvae was measured every four to five days until adult eclosion. The cumulative proportion dead was transformed by the arcsine square root transformation and treatments were compared with ANOVA. Means were separated with Duncan’s Multiple Range test. To examine the temporal pattern of mortality, the mean cumulative mortality per tree was plotted against days post-injection, and nonlinear regression used to fit curves to data.
Presence of B. t. antigens was also investigated by immunoassays. Leaves and larvae in the B.t. treatment and controls were collected and frozen. Samples of leaves and larval midguts were tested for B.t. antigens using an ELISA for the leaves and a western blot test for the midguts by Dr. R.A. Smith, Abbott Laboratories.
Results
The NPV and acephate treatments were applied with no difficulty, but the trees did not take up the B.t. suspensions as easily. Four liters of NPV suspension was taken up by the trees in a mean (±SE) of 60 ±25 minutes per tree. In contrast, we were unable to put the entire four liters of B. t. suspension into any of the three B.f. replicates, and there was a great deal of foaming of the suspension during injection. The first B.t. tree took ca. 3.2 liters in 3.5 hours, the second ca. 2.4 liters in 5.5 hours, and the last replicate 2.2 liters in 3.5 hours.
Efficacy
Trees treated with acephate showed a significant difference in the proportion dead (Table 1) when compared to any of the other treatments. The cumulative mortality from the acephate treatment was higher (P < 0.05) than controls and all other treatments 20 days post application and thereafter. No significant differences were observed among any of the other treatments, including controls. All mortality in all treatments occurred only in the larval stage. With the exception of one vestigially winged adult in the NPV treatment, no genetic or growth abnormality was observed from any treatment.
Mean ± SE cumulative percent mortality of gypsy moths reared on foliage from pin oak trees treated with synthetic organic and microbial insecticides.1
Temporal pattern of mortality
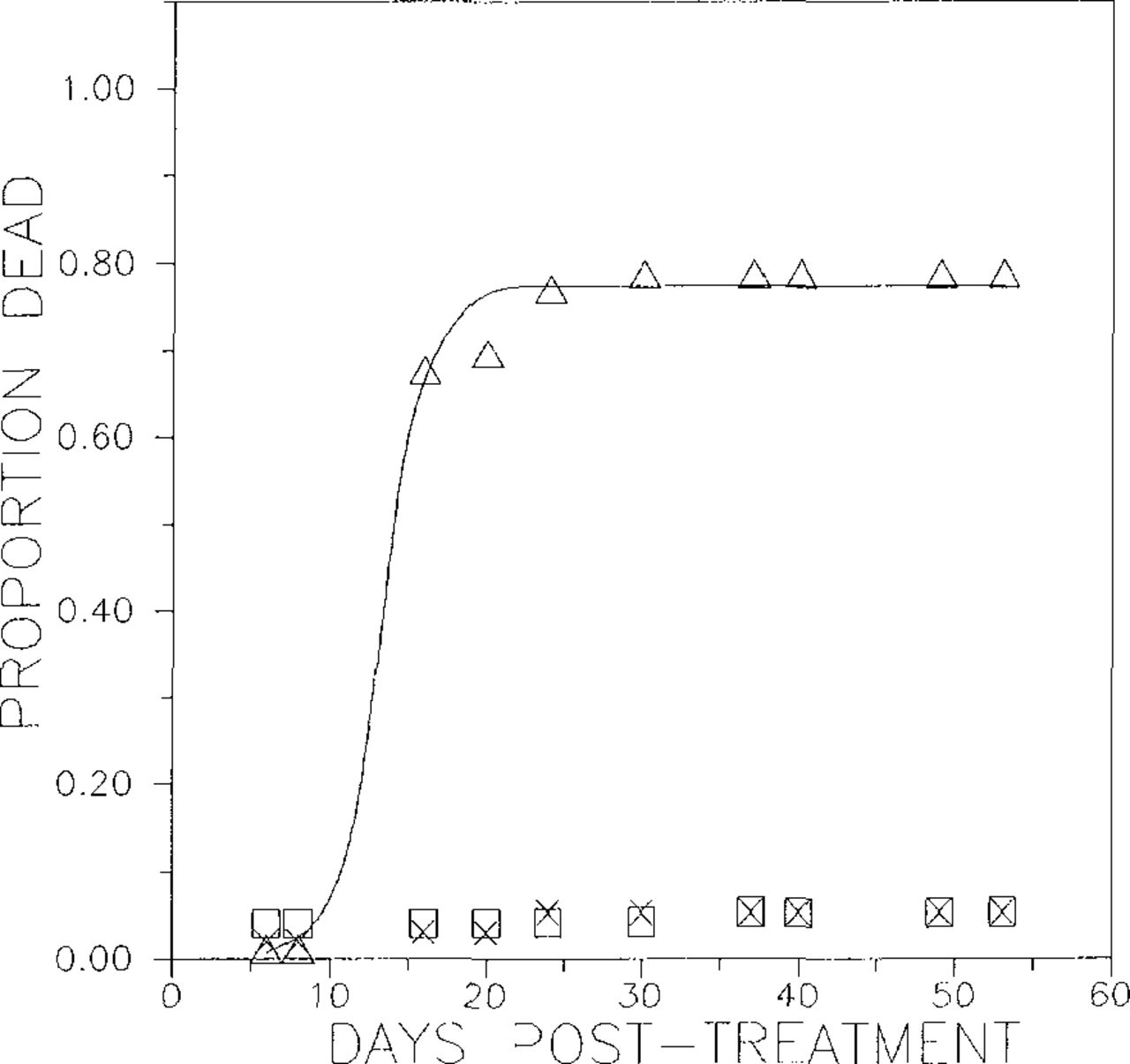
The pattern of mortality over time was distinctive but highly variable among trees. In the acephate treatment, cumulative mortality over time followed an exponential sigmoid model (R2 = 0.76, Fig. 1). There was considerable variation among trees in the length of lag time prior to showing a response. Two replicates passed the lag phase and showed a strong response by the first time mortality was measured, 6 days post treatment, whereas the third tree did not show this response until ca. 18 days post treatment. Most of the residual variance in the model was due to differences in lag time among tree replicates. The 95% confidence interval for the intercept (B in the model presented in the figure caption) overlapped zero, suggesting that the data reasonably represent no mortality prior to treatment. All other regression parameters were significant (i.e., they had 95% confidence intervals that did not overlap zero).

Temporal pattern of mortality of gypsy moths reared on foliage from three trees treated with acephate implants. Each point represents the mean cumulative proportion mortality from five cages; data from different trees are plotted with different symbols. Data from three trees fit to an exponential sigmoid pattern y = Pmax/(1 + Benx), where y Is the cumulative proportion mortality and x is days post treatment. Regression coefficients (± SE) are Pmax = 1.01 ± 0.05; B = 20.67 ± 16.66; and n = -0.22 ± 0.06.
In the B.t. trees, two replicates had no mortality in excess of that observed from control trees, but one replicate followed the exponential sigmoid pattern of cumulative mortality over time (R2 = 0.99, Fig. 2). No antigen was detected, however, in any leaf or larval midgut samples, which included dead larvae from the one replicate that showed a bioassay response. The high R2 is due to the lack of tree replications used to fit the model. As in the acephate treatment, all regression coefficients were significant except the one expressing mortality at time 0 (the B parameter). No regression models were developed from the control or NPV trees due to the low rate of mortality throughout the experiment (mean ± SE at 51 days post injection of 11 ±4 and 7 ± 2 in the control and NPV trees, respectively).
Temporal pattern of mortality of gypsy moths reared on foliage from trees treated with pressurized Injection of Bacillus thurlnglensls var. kurstaki. Each point represents the mean cumulative proportion mortality from five cages; data from different trees are plotted with different symbols. Data from the one tree that showed a response fit to an exponential sigmoid pattern y = Pmax/ (1 + Benx), where y Is the cumulative proportion mortality and x Is days post treatment. Regression coefficients (± SE) are Pmax = 0.78 ± 0.01; B = 5556 ± 12632; and n = -0.65 + 0.15.
Discussion
This study supports the work of Webb et al. (10) on the feasibility of using acephate implants (ACECAPS) for controlling gypsy moths on individual trees. The material in the implant formulation and the application procedure is approved for use by individual homeowners and is easy to implement. The time to achieve mortality suggests that implantation be applied early if leaf feeding is to be minimized. Webb et al. (10) inserted implants around the time of budswell to budburst, and achieved 50% mortality in larvae exposed to treated foliage for 4-day intervals. Our data show the higher rates of mortality that can be expected and the time to achieve these rates when larvae feed continuously on foliage from the treated tree. However, other differences in the experimental designs may have caused the differences in mortality rates.
Further efforts are needed to explain why NPV gave no significant mortality and B.t. gave variable results. The pH of the sap on April 14 ranged from 5.0 to 5.2. Some microbial insecticides are activated by partial digestion in the alkali environment of the lepidopteran gut. The acidic pH of the tree sap suggested that the microbial insecticide should not be adversely affected during ascent in the sap stream. Foaming of the B.t. suspension may be reduced by incorporation of a surfactant, and this may result in more of the material going into the tree. It is also conceivable that the rotary knife blender may have physically disrupted the insecticidal crystals of both microbial treatments. Homogenization of the mosquito larvicide B.t. var. israelensis with a rotary knife blender has been shown to reduce efficacy (9). Another hypothesis is that the lack of efficacy could be due to polymerization of proteins of the NPV coat or the B.t. spore due to binding with tannins. In addition to further work with the microbials, other materials that show promise for environmentally-sound control of gypsy moths should be tested for implantation or injection application techniques.
Implantation or injection of insecticides will undoubtedly have a place in integrated pest management programs directed against the gypsy moth in the urban environment. As with any insecticide, care must be taken to avoid abuse. The formulation involved in implantation involves a high proportion of active ingredient relative to formulations most often applied by homeowners. Any insecticidal treatment should only be applied after sampling to estimate insect density and relating that density to management objectives.
Acknowledgments
We sincerely appreciate the ACECAPS for injection supplied by Creative Sales, Inc.; the B.t. from R.A. Smith at Abbott Laboratories; and the NPV from J.D. Podgwaite, USDA-FS, NE Forest Insect and Disease Laboratory. Dr. Smith and Dr. Podgwaite also made the cleaned formulations for injection. P. Parkman, D. Gray, and K. Eaton helped with the bioassay when the authors were unavailable. This research was funded by a competitive grant from the International Society of Arboriculture Research Trust Fund and Forest Pest Management, USDA Forest Service, Morgantown, WV.
Footnotes
↵1 Department of Plant Pathology, Physiology and Weed Science.
- © 1989, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.