
Abstract
Root collars of 11-yr-old pin oak (Quercus palustris), red maple (Acer rubrum), white ash (Fraxinus americana), and black walnut (Juglans nigra) were drenched in June, August, or October 1985 with 2 or 4 g a.i. (W.P.) flurprimidol {a-(1-methylethyl)-a-[4-(trifluoromethoxy)-phenyl]-5-pyrimidine-methanol) to determine physiological responses. No treatment response occurred the year of treatment but significant terminal and lateral growth inhibition, 43 to 92%, respectively, occurred during the first growing season (1986) following treatment. In general, less growth inhibition resulted from October treatments. However, by the end of the second growing season following application (1987), all treatment combinations inhibited terminal and lateral growth. Leaf area of pin oak and red maple only was reduced (22 to 65%) in 1986 but by 1987 leaf area was reduced 17 to 65% on all species from all treatments. Except for white ash, leaf dry weight was reduced (28 to 64%) by 1987. In general, flurprimidol caused a slight decrease in photosynthetic activity and an increase in chlorophyll concentration but transpiration rates were unchanged. These results indicate that flurprimidol reduces growth without significantly altering energy assimilation or water movement in individual leaves.
Urban foresters are investigating the use of growth inhibitors to lower the high cost of mechanical tree trimming by reducing the need for pruning. Flurprimidol is one of a group of gibberellin biosynthesis inhibitors including paclobutrazol and uniconazole. They offer the potential for use as tree shoot elongation inhibitors and they are of particular interest because they are not phytotoxic to woody plants and may be applied to roots, buds, or vascular systems (3, 5, 8, 9, 11, and 12). In other studies, flurprimidol applied as a root collar drench in early spring in Maryland uniformly suppressed the vegetative growth of white ash (Fraxinus americana) and black walnut (Juglans nigra) the second growing season after treatment (10) and when applied as a subsoil injection it also suppressed the growth of red maple (Acer rubrum) seedlings in Alabama (4). Most photosynthesis research with woody plants using gibberellin biosynthesis inhibitors has been conducted on fruit trees with paclobutrazol. Steffens and Jacobs (7) reported that photosynthetic rates from paclobutrazol-treated apple trees were not changed but photosynthesis on a per tree basis was reduced because total leaf area was reduced and leaf stomatal resistance showed no consistent pattern. Wieland and Wample (13) reported similar results on apple trees. This investigation was designed to determine physiological responses of four deciduous shade trees treated during three different growth stages with root collar drenches of flurprimidol.
Materials and Methods
Growth response
Eleven-year-old pin oak (Quercus palustris), red maple, white ash, and black walnut saplings (5 m to 8 m tall; 7 cm to 8 cm diameter) were grown on 3 m × 3 m centers in Duffield silt loam in Frederick, MD. Root collars of these trees were drenched in 1985 with 250 ml of 50% wettable powder formulation of flurprimidol (2 or 4 g a.i. per tree) during (a) rapid growth in June, (b) reduced growth in August, or (c) the onset of dormancy in October. The soil at the tree base was pushed back about 2 cm with a trowel to expose the root collar bark. Crown development was evaluated during 1986 and 1987 in terms of terminal leader and lateral branch growth; leaf area, dry weight and rachis length; and diameter breast height (dbh) of the main stem. Terminal leader growth was determined by measuring the height of each tree before treatment and at the end of each growing season. Lateral growth was determined by measuring the current years shoot growth from 4 lateral branches located low in the crown radiating approximately north, east, south, and west from the trunk. These branches were marked with flagging before growth occurred. Leaf area, dry weight, and rachis length of species with compound leaves were determined near the end of the 1986 and 1987 growing seasons from the three largest leaves nearest the shoot tip of a lateral branch (4 lateral branches = 12 leaves per tree). Leaf area was measured with a photoelectric leaf-area meter (Lambda Instruments). Trunk diameter growth was determined with calipers before treatment and at the end of the 1987 growing season. Treatments were replicated 5 times using a randomized complete block design. Means were separated by Waller-Duncan K-ratio t test.
Photosynthetic response
In August 1987 when inhibition was obvious, net photosynthesis (Pn), transpiration, and chlorophyll content were determined on untreated pin oak, red maple, white ash, and black walnut leaves and on leaves of trees treated in June 1985 with 4 g flurprimidol (the most uniformly effective treatment). Pn and leaf transpiration were measured simultaneously on four sunny days from 9:00 AM to 11:30 AM (photosynthetically active radiation ranged from 800 to 1800 uE s_1 m~2) using a field measuring technique (LiCor 6000 Portable Photosynthesis System). Two fully expanded leaves located near the shoot tips and exposed to full sunlight were removed from the upper part of the crown and two from the lower part of the crown on 4 trees per species per day. Each leaf was measured immediately after removal for Pn and transpiration. These same leaves were refrigerated in the field and analyzed in the laboratory for chlorophyll content (2). At least three replications (1 tree/replicate) were made, the values of the four leaves per tree were averaged and standard error of the mean determined.
Results and Discussion
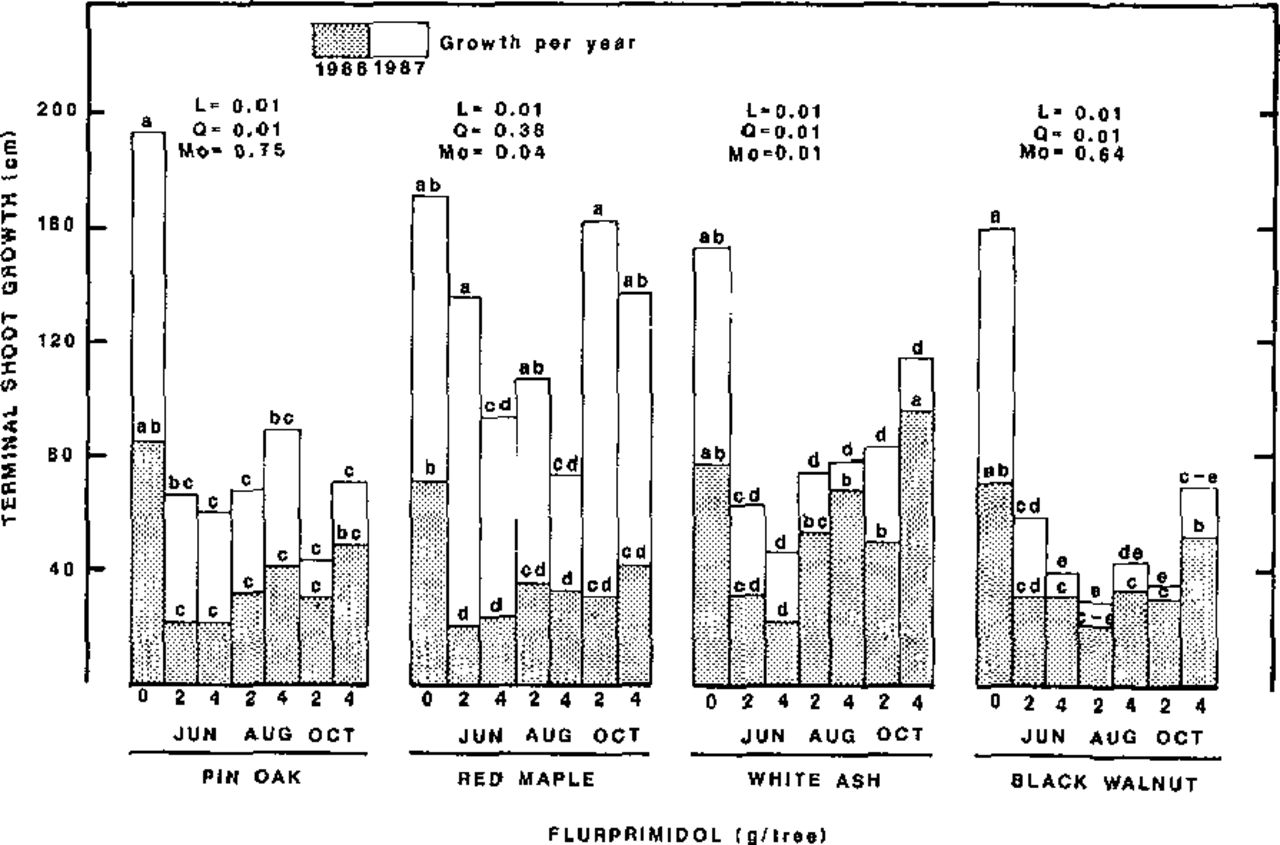
The effect of 1985 flurprimidol root collar drenches was not evident until mid-June 1986. Terminal growth was inhibited (43 to 70%) on all species-treatment-time combinations by the end of 1986 except for August and October-treated white ash (Figure 1). In 1986 lateral shoot growth of pin oak and red maple was inhibited by June and August treatments (50 to 92%) but the June treatment was most effective for white ash in 1986 (Figure 2). Lateral growth of black walnut in 1986 was unaffected by flurprimidol. Increasing the level of flurprimidol from 2 to 4 g per tree did not increase inhibition at the same rate as 0 to 2 g per tree. This is indicated by a significant linear and quadratic component to the regression equations. Terminal shoot growth in 1986 of October treatments on ash and walnut was significantly more from 4 g versus 2 g flurprimidol (Figure 1). A possible explanation for this anomaly is that root absorption was slowed in the presence of the high concentration of flurprimidol (16,000 ppm versus 8,000 ppm in water) thus slowing transport to the terminal shoot apices. However, by the end of the 1987 growing season inhibition from both rates was not different.

Terminal shoot growth at the end of the second and third growing seasons following root collar treatment of 11-year-old trees with flurprimidol in June, August, and October 1985. Within species, means are separated with Waller-Duncan K-ratio t test, 5% level. Contrasts above histograms display probability values of linear (L) or quadratic (Q) effects due to flurprimidol rate and to month of application effect (Mo).

Lateral growth per shoot at the end of the second and third growing seasons following root collar treatment of 11-year-old trees with flurprimidol in June, August, and October 1985. Within species, means are separated with Waller-Duncan K-ratio t test, 5% level. Contrasts above histograms display probability values of linear (L) or quadratic (Q) effects due to flurprimidol rate and to month of application effect (Mo).
By the end of the 1987 growing season terminal and lateral growth of all species was significantly inhibited by all six treatments (33 to 88%) except for terminal shoot growth of red maple from October treatments and the 2-g rate in June and August treatments (Figures 1 and 2). Red maple was the first to show obvious signs of inhibition in 1986 but was the only species to regain normal terminal growth in 1987; although, the 4-g rate applied in June still effectively inhibited terminal shoot growth of red maple in 1987. Excavations around the root collars of the four species treated revealed that red maple had a much higher number of nonwoody (fibrous) roots growing from older woody roots in the vicinity of the root collar zone than the other three species. This may explain in part, red maples’ faster inhibition response to flurprimidol. Although not significant at the 5% level due to variability among replications, there appeared to be some inhibition of diameter growth (data not shown).
Leaf area in 1986 was reduced for pin oak, red maple, and white ash (22 to 65%), but not for black walnut. In 1987 leaf area was inhibited (17 to 65%) for all species treatment combinations (Table 1). Leaf rachis length in 1986 was reduced by June and August treatments on white ash (20 to 45%) resulting in a more compact leaf. Rachis length of black walnut was not affected in 1986, but in 1987 rachis length was signficantly reduced (P ► 0.01) for both white ash and black walnut 21 to 50% (data not shown). In 1986, leaf dry weight of red maple treated in June and August differed from controls, but leaf dry weight in 1987 was reduced for all species (28 to 64%) with the exception of white ash (Table 2). Increasing flurprimidol level from 2 to 4 g per tree did not decrease leaf area or dry weight at the same rate as 0 to 2 g per tree since there were signficant linear and quadratic components in the regression equations (Tables 1 and 2).
Leaf area of 11-year-old trees that were root-collar drenched with flurprimidol in 1985.
Leaf dry weight of 11-year-old trees that were root-collar drenched with flurprimidol in 1985.
With the exception of pin oak terminal growth, less inhibition of growth and leaf expansion occurred from the October root collar drenches than June or August as indicated by the significant month effect in Figures 1 and 2, and Table 1. The less effective inhibition in October was probably because the trees were not actively translocating acropetally at treatment time and had one less growing season to absorb and translocate flurprimidol than trees treated in June or August. Also, no significant effect on growth was observed on any species-treatment-time combination during the growing season (1985) the root collar drenches were performed. Since trunk-injected trees usually respond to flurprimidol within a month or two after treatment (9), time of absorption through the older root bark must be responsible for the slow inhibition response. However, by the end of the 1987 growing season, shoot growth inhibition of both terminal and laterals was both effective and very uniform without apparent shoot “escapes” except for red maple treated with the lower rate. Gilliam et al. (4) observed that red maple “escape” shoots were even longer than control tree shoots three years after treatment of seedlings. The “escape” shoots of treated trees in this study after three years were not significantly longer than control trees.
In addition to the reduction in leaf area and dry weight, flurprimidol caused a slight decrease in net photosynthesis and an increase in chlorophyll concentration (Table 3). The transpiration rate was not affected. Steffens and Jacobs (7) also found a lower relative photosynthetic rate of apple trees with the gibberllin biosynthesis inhibitor paclobutrazol. One would expect leaf photosynthetic rates to be higher than control trees since chlorophyll concentration was higher. However, Anderson (1) found that the photosynthetic rate under natural conditions does not vary much over a considerable range of chlorophyll concentrations and that the rate of photosynthesis is usually well below the maximum possible rate (6). In this study, the reduction in the photosynthetic rate, leaf dry weight and expansion that coincided with the increased chlorophyll concentration did not appear to adversely affect plant vigor or aesthetic quality. The only visible symptoms of flurprimidol in 1986 and 1987 were small, dark-green leaves and shorter shoots. In an earlier study, white ash and black walnut trees treated in April 1984 with a dose as high as 12 g/tree of flurprimidol were still visibly healthy in June 1988 with obviously high concentrations of leaf chlorophyll and shorter shoots (10).
Physiological responses of 11-year-old trees to a root-collar treatment of flurprimidol (4 g/tree, June 1985). Measurements were made in August 1987.
Conclusions
The results of this study indicate that root collar treatments of flurprimidol conducted during the growing or early dormant season inhibited growth uniformly without visible phytotoxicity for two years and, with the exception of growth inhibition, had a relatively minor effect on physiological processes. In addition to consistent uniform inhibition of terminal and lateral shoot growth, which is difficult to obtain with trunk injection techniques, the root collar drench method has several advantages especially for controlling the growth of young trees. Root collar zones of city trees are usually accessible whereas the fibrous roots (targets for the soil injection technique) that extend beyond the foliage canopy may be unavailable; e.g., covered with concrete or macadam. Confining flurprimidol to the root collar zone reduces the possibility of contacting nontarget plants because flurprimidol remains close to the root collar zone. In another study, using a bioassay technique, flurprimidol activity was still confined laterally within 10 cm of the treated root collar 1 ½ years after drenching the base of the trunk (10). Before flurprimidol can be registered for root collar drench use, more information is needed regarding effective doses for large trees and a variety of other species. Also, information is needed regarding longevity and movement of furprimidol residues in various soil types. When drenching the root collar zone, some of the wettable powder formulation, although directed at the trunk base, makes contact with the soil.
Acknowledgments
We gratefully acknowledge the technical assistance of N. Monath and E. Johnson, and we thank Lilly Research Laboratories for furnishing samples of flurprimidol (Cutless, EL5OO). The authors also thank J.G. Phillips for statistical assistance.
- © 1989, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.