
Abstract
Effects of root-zone temperatures (28°, 35° and 42°C; 6 hrs/day for 12 weeks) on container-grown ‘East Palatka’ holly (llex x attenuate ‘East Palatka’) and ‘Drake’ elm (Ulmus parvifolia ‘Drake’) were determined under growth-room conditions. Survival rate of elm was decreased by 50% after 12 weeks of daily exposure to 42°C. Temperature did not affect shoot to root ratios of either genus. However, dry weights of holly shoots and roots and elm roots decreased quadratically, and elm shoots decreased linearly with increased temperature. Time trend analysis for holly and elm shoot extension, leaf carbon exchange rates (CER), and stomatai conductance (Cs) revealed reductions at 42°C compared to 35° and 28°C for both genera. Results demonstrate the need for tree growers to execute cultural practices directed at minimizing exposure of tree roots to temperatures at or above 42°C.
Temperature is a major factor affecting plant growth. Although surface soil temperatures have been observed to surpass critical limits resulting in physiological disorders such as “strangulation sickness” in pine seedlings (12), the outstanding examples of high temperature exposure to root systems are in nursery container media exposed to direct solar radiation. Average medium temperatures during the summer may exceed 40°C for up to 6 HR daily (6), and a mean maximum temperature of 58°C has been recorded on the western exposure (15).
Elevated root-zone temperature below a critical killing threshold can alter plant source-sink relationships and thus carbon partitioning. Langridge and McWilliam (11) suggested that temperature becomes increasingly important as a determinant in the survival distribution of plants the farther a plant deviates from its biological norm. Carbon partitioning to shoots and roots in response to root temperature is species dependent (8).
The effects of air temperature on photosynthesis and carbon exchange rates have been studied for many plants (1, 2). However, root temperature also alters photosynthesis of selected species (3,5). Leaf photosynthetic rates of Pittosporum tobiria were lowered at 40°C root temperature compared to 27° and 32°C (9).
‘East Palatka’ holly and ‘Drake’ elm are landscape trees frequently grown in containers in the southern United States. Container culture of trees generally require extended periods compared to shrubs, potentially increasing the number of exposures of container walls to insolation. Research was conducted to ascertain the growth and physiological response of ‘East Palatka’ holly and ‘Drake’ elm to three root-zone temperatures.
Materials and Methods
Eight- to 12-cm rooted cuttings of Ilex x attenuate ‘East Palatka’ and Ulmus parvifolia ‘Drake’ were planted individually into polyethylene sleeves (26 × 7.5 cm in diameter) using a Metro-Mix 500 growth medium (W.R. Grace and Co., Cambridge, MA.) during the fall of 1987. Trees were allowed a 3-month growth period in a temperature-controlled glass greenhouse (37°/17°C) under natural daylength supplemented with 4 hr of incandescent light to maintain active growth.
A 12-week experiment was conducted in a 3 × 7.5 m walk-in growth room with irradiance supplied by eighteen 1000 wait phosphor-coated metal-arc HID lamps (GTE Sylvania Corp., Manchester, NH). Light intensity measured by a quantum radiometer (LI-COR Inc., Lincoln, NE) at plant height was 1150 μmol−2s−1. Photoperiod was maintained at 13 hours (0815 to 2115 HR) supplemented with a 3 hour incandescent light during the dark period. Air temperature was 28° to 3O°C/day gradually changing to 20 to 22°C/night. Relative humidity was approximately 40%/90% (day/night) as recorded with a hygrothermograph. Root temperatures of 28°, 35°, and 42°± 0.8°C were sustained 6 hours per day, 1000 to 1600 HR, using an electronically controlled root-heating system (4). Polyethylene sleeves were placed into galvanized metal tubes (23 × 7.5 cm in diameter) wrapped with 60 watt 120 vac heating tape (Smith-Gates Corp., Farmington, CT) and 1,25-cm thick insulation. These root heating tubes were connected to solid state electronic controllers which regulated preset temperatures via a thermistor feedback mechanism.
Shoot length from a preset point of origin was measured at 2 week intervals during the experiment. Shoot extension was then calculated based upon initial shoot length measurements. At weeks 1,4, 8, and 12, CER and Cs were measured using a portable photosynthesis system (LI-6000 model, LI-COR, Inc., Lincoln NE). The most recently expanded leaf was sampled for each tree (typically 3 to 5 leaves from the shoot apex). A 0.25 liter cuvette chamber was used for measurements and the mean of 10 consecutive observations on each leaf comprised an experimental unit. Leaf area for holly was determined by partitioning the cuvette chamber surface into a known surface area and estimating the area of a recently matured leaf. For elm, leaf area (LA) was approximated using a leaf area model based upon maximum leaf width (W). Regression analysis was used to develop the model equation LA = 1,73W2 − 1.68W + 2.09 (r2 = 0.98). At week 12, the experiment was terminated when roots were severed from shoots and thoroughly washed. Roots and shoots were then oven dried at 70°C for 48 hours and dry weights were determined.
The experimental design was a completely randomized design with 10 single tree replicates. Regression analysis (Statistical Analysis System, Raleigh N.C.) was used to test for significant linear and quadratic responses of the variables to temperature treatments at week 12. Regression coefficients were tested for homogeneity of fit using the F test. Variable responses over time were tested by repeated measures analysis. Probabilities for the F test for time and time x temperature interactions were made using the Greenhouse-Geiser adjustment to univariate P values (13).
Results and Discussion
Holly
Root and shoot dry weights decreased quadratically with increased root-zone temperature (Table 1). Root and shoot dry weights were reduced 78% and 73%, respectively, at 42°C compared to 35°C.
Effect of root-zone temperature on dry weights and shoot to root ratio (S/R) of East Palatka holly and Drake elm.
Root-zone temperature interacted with exposure time to affect all measured variables. Time trends for shoot extension at 28°C and 35°C were similar and increased linearly compared to 42°C (Figure 1a). Hollies grown at the 42°C treatment were generally stunted in appearance with small, yellowish-green foliage. Leaf CER and Cs decreased for all treatments at week 4 compared to week 1 indicating that holly may require more than 4 weeks to acclimate to growth room conditions (Figure 2a and 2b). Leaf CER and Cs at 35°C and 28°C increased linearly from weeks 4 to 12. However, the rate of increase for leaf CER at 28°C was greater than at 35°C. The 4 to 12 week time trends for leaf CER and Cs at 42°C were quadratic compared to 35°C and 28°C as evidenced by the recovery in leaf CER and Cs at weeks 8 and 12 compared to week 4. Recovery rate at 42°C was greater for leaf CER compared to Cs, suggesting that photosynthetic efficiency as a function of chlorophyll enzymatic activity may increase over time and that regulation of ‘East Palatka’ holly photosynthesis may be in part non-stomatal. This conclusion supports earlier findings by Foster et al (4) who suggested a non-stomatal inhibition of photosynthesis in Ilex crenata ‘Rotun-difolia’ exposed to supraoptimal root-zone temperature.

Effect of root-zone temperature on shoot extension of ‘East Palatka’ holly (1a) and ‘Drake’ elm (1b).
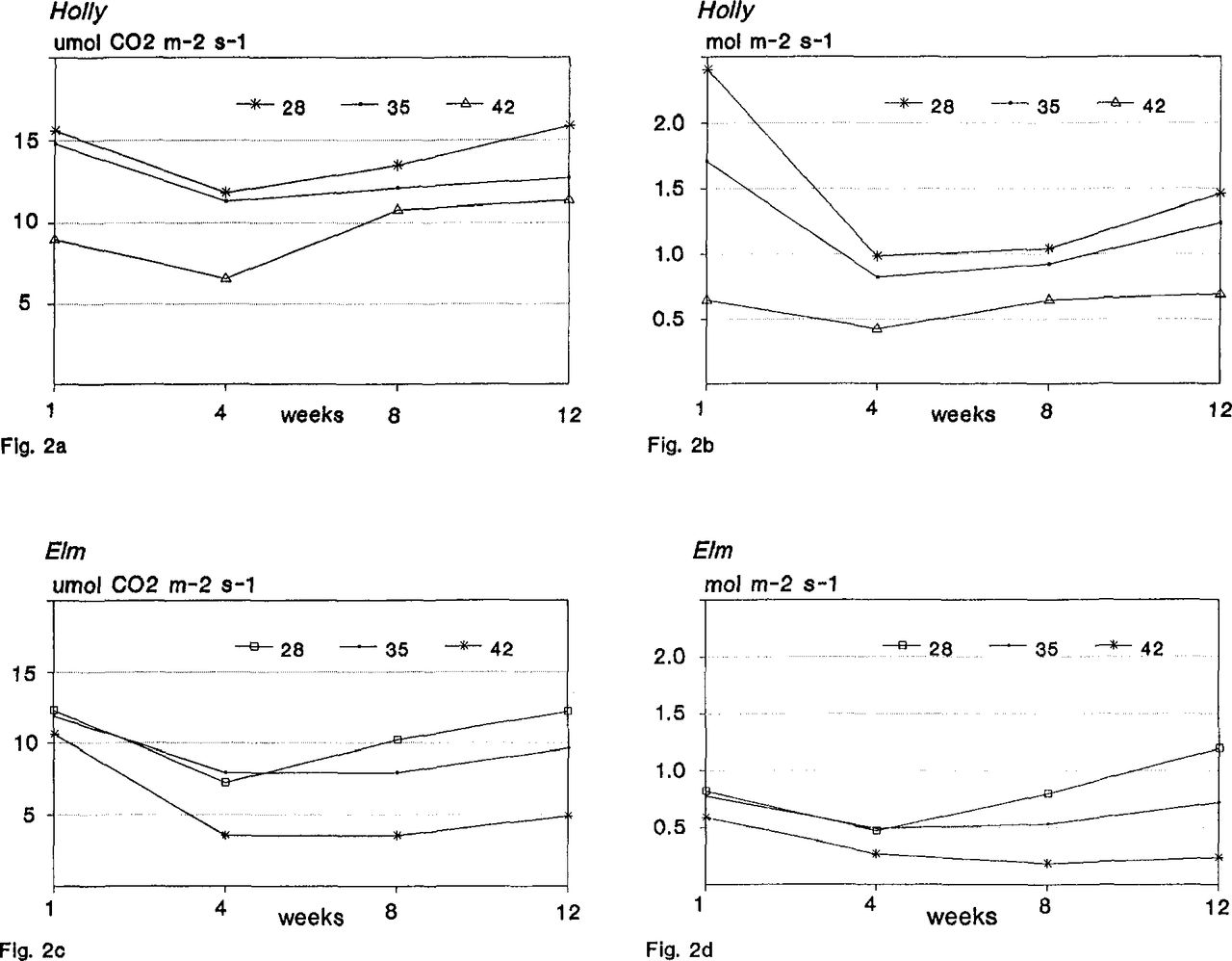
Effect of root-zone temperature on leaf carbon exchange rates and stomatal conductance of ‘East Palatka’ holly (2a& 2b) and ‘Drake’ elm (2c& 2d).
Elm
Survival rate was affected by root-zone temperature. Although all trees exposed to 28°C and 35°C survived, 50% of trees at 42°C had died by week 12. Shoot dry weights decreased linearly and root dry weights decreased quadratically with increased root-zone and temperature (Table 1). Shoot and root dry weights at 42°C were reduced by 78% and 68%, respectively, compared to 35°C.
Root-zone temperature interacted with exposure time to affect all measured variables. Time trends for shoot extension at 28°C and 35°C were similar and increased linearly compared to 42°C (Figure 1b). Although shoot extension after 12 weeks at 35°C was 4% greater than at 28°C, elms grown at 28°C were observed to have more lateral branches. Similar to holly, leaf CER and Cs decreased for all treatments at week 4 compared to week 1 suggesting that elm may require greater than 4 weeks to acclimate to growth room conditions (Figure 2c and 2d). Leaf CER and Cs at 35°C and 28°C increased linearly from week 4 to 12; however the rate of increase at 28°C was greater than at 35°C. Leaf drop of older leaves, a common response of deciduous trees to environmental stress (10), was observed after week 4 for the 42°C treatment (data not shown). The fact that leaf drop and reduced CER and Cs occurred after week 4 with little to no recovery in rates suggests that 1) 42°C is above the temperature compensation point where photosynthetic and respiratory rates are equal and, 2) elm was not able to acclimate to treatment temperature over a 12 week period.
The optimum temperature treatment for tree growth during this time period as suggested by dry weight accumulation, appeared to be 35°C for holly and 28°C for elm with significant reductions in biomass accumulation occurring in trees at 42°C. The shoot to root ratio was not affected by root-zone temperature and averaged 4.5 for holly and 1.8 for elm (Table 1). Root-zone temperature has altered the shoot to root ratio of other woody plants but whether the ratio was increased or decreased was species dependent (6, 8, 9).
The measured physiological responses differed according to genus. Decreased CER and Cs for elm at 42°C without a recovery in rates within 12 weeks, indicates that supraoptimal root-zone temperature may affect the aperture of stomatai guard cells thus regulating photosynthesis. Stomatal aperture also controls the rate of water loss and is considered to be in part regulated hormonally by abscisic acid and root-synthesized cytokinins (14, 16). Damage to roots reduces the capacity for water uptake. For elm at elevated root-zone temperature, the control of water loss (transpiration data not shown) may govern the regulation of stomatal aperture. In contrast, the recovery of leaf CER over time at 42°C supports the hypothesis that holly may partially acclimate to supraoptimal root-zone temperature.
In summary, this study indicates that ‘East Palatka’ holly tolerates higher root-zone temperature than ‘Drake’ elm; however, the growth of both genera will be reduced by rootzone temperatures typically observed in southern nurseries. Reduced tree vigor induced by high temperature exposure during production may result in increased production time and unit costs in addition to reduced tree survival and establishment rates upon transplanting into the landscape. Container spacing strategies that minimize the exposure of nursery container walls to direct solar radiation should be considered by nursery operators as a means of minimizing root-zone temperature fluxes (7).
Footnotes
Received for publication 6 June 89. Florida Ag. Expt. Sta. J. Ser. No. 9948.
- © 1989, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.