
Abstract
Two experiments were conducted to determine the effects of moisture stress on one-year-old containerized Cornus florida. Recovery times of plant’s shoot water potential, stomatal conductance and transpiration to the levels of nonstressed control plants were dependent on duration of moisture stress. Increased stress resulted in longer recovery periods. Shoot length was suppressed by severe moisture stress. Short periods of moisture stress, such as 10 days without water followed by normal watering, resulted in initial suppression of shoot length, but by the end of the growing season, there were no differences between short-term moisture-stressed plants and nonstressed plants. Longer periods of stress, such as 22 days without water, resulted in suppressed shoot growth in the growing season of the stress and also produced a slower shoot growth rate the following growing season.
Flowering dogwood is widely grown in the Southeastern United States. When dogwood is subjected to drought, vegetative growth is affected (1, 3, 4), flower and bud formation may be suppressed (3), and marginal leaf scorch may occur (3). Kozlowski and Davies (2) point out that even mild periods of moisture stress can be detrimental to plants. Kozlowski (3) identified 3 stages of wilt: incipient wilt - a slight decrease in turgor during the day; temporary wilt - visible drooping of leaves during the day but recovery of turgor overnight; and permanent wilt - plants continue in a wilted condition ending in death unless water is added.
Periods of drought stress occur almost annually in the Southeast. As a result, management techniques designed to alleviate or reduce moisture stress are needed. To develop these management techniques, basic information is needed on the response of dogwood to moisture stress.
Materials and Methods
Experiment 1
In January, 1984, 240 one-year-old bareroot Cornus florida liners were potted in 11.4-liter (#3) containers. The potting medium consisted of pine bark and sand (4:1 by volume) amended on a m3 basis with 3.5 kg dolomite, 1.2 kg gypsum, 1.2 kg superphosphate, 0.6 kg Aquagro, 0.9 kg Micromax, and 8.3 kg of 18N-2.5P-1 OK (18-6-12) Osmocote.
Plants were maintained in full sun with overhead irrigation until 5 June 1984, when 24 plants were relocated to raised benches in a double layer polyethylene greenhouse where temperatures ranged from 20°C to 32°C during the experiment. One week later the following treatments were initiated: 1) plants watered as needed (control); 2) plants watered only when they reached the permanent wilting point (plants wilted during the day and did not recover overnight); and 3) plants not watered until at least half of the plants exhibited leaf necrosis. After plants reached the above indicated physiological stages, they were watered and maintained in the same manner as the control plants through the experiment (30 June, 1984). The experimental design was a completely randomized design with 6 single plant replicates.
Data taken included shoot water potential, stomatal conductance, and transpiration. Shoot water potential was measured in bars using a Scholander Pressure Bomb (PMS Instrument Company, Corvallis, Oregon). Stomatal conductance and transpiration were measured in mmol m−2s−1 using a LiCor LI 1600 Steady State porometer (LiCor Incorporated, Lincoln, Nebraska). Data were taken daily at 7:00 a.m., 11:00 a.m., 3:00 p.m., and 7:00 p.m. until day 8,at which time the 7:00 p.m. measurements were discontinued.
Experiment 2
On 27 June, 1984, a group of 80 one-year-old uniform Cornus florida similar to those in Expt. 1 were moved from the outside nursery area to a polyethylene greenhouse. Greenhouse temperatures ranged from 20°C to 32°C.
One week later the following treatments were initiated: 1) watered as needed (control); 2) no water until day 5 (plants reached permanent wilting point), then watered as needed afterward; 3) no water until day 5, watered once then restressed by withholding water until day 18, and thereafter maintained similarly to the watered controls; 4) no water until day 10 (half of the plants exhibiting leaf necrosis), and thereafter maintained similarly to the watered controls; 5) no water until day 10, watered once, restressed until day 18, and thereafter maintained similarly to the watered controls; 6) no water until day 22 (severe necrosis and twig dieback), and thereafter maintained similarly to the watered controls; 7) no water until day 38, then watered similarly to the watered control plants. Water was applied about 3:00 p.m. for all treatments on the specified days.
In this experiment 3 shoots per plant were randomly selected and measured weekly from 2 July to 4 November when plants were placed outside for overwintering. On 31 March, 1985, these plants were placed in the greenhouse and weekly shoot length measurements were resumed. Measurements were terminated on 2 May, 1985 (the end of the first growth flush).
Plants were arranged in completely randomized design with 10, two-plant replicates of the watered control and 5, two-plant replicates of the other treatments.
Results and Discussion
Experiment 1
Shoot water potential of the watered plants ranged from −4 to −12 bars from days 6-18 (Fig. 1): transpiration ranged from 2.4 to 8.4 mmol m−2s−1 (Fig. 2), and stomatal conductance ranged from 140.5 to 253.5 mmol m−2s−1 (Fig. 3). These plants did not show loss of turgor or exhibit leaf necrosis.

Shoot water potential of dogwoods watered day 8, dogwoods watered day 12 and watered control in Experiment 1; measured daily at 7:00 a.m., 11:00 a.m., and 3:00 p.m. beginning 12 June, 1984. Mean separation is by date using Duncan’s multiple range test, 5% level.
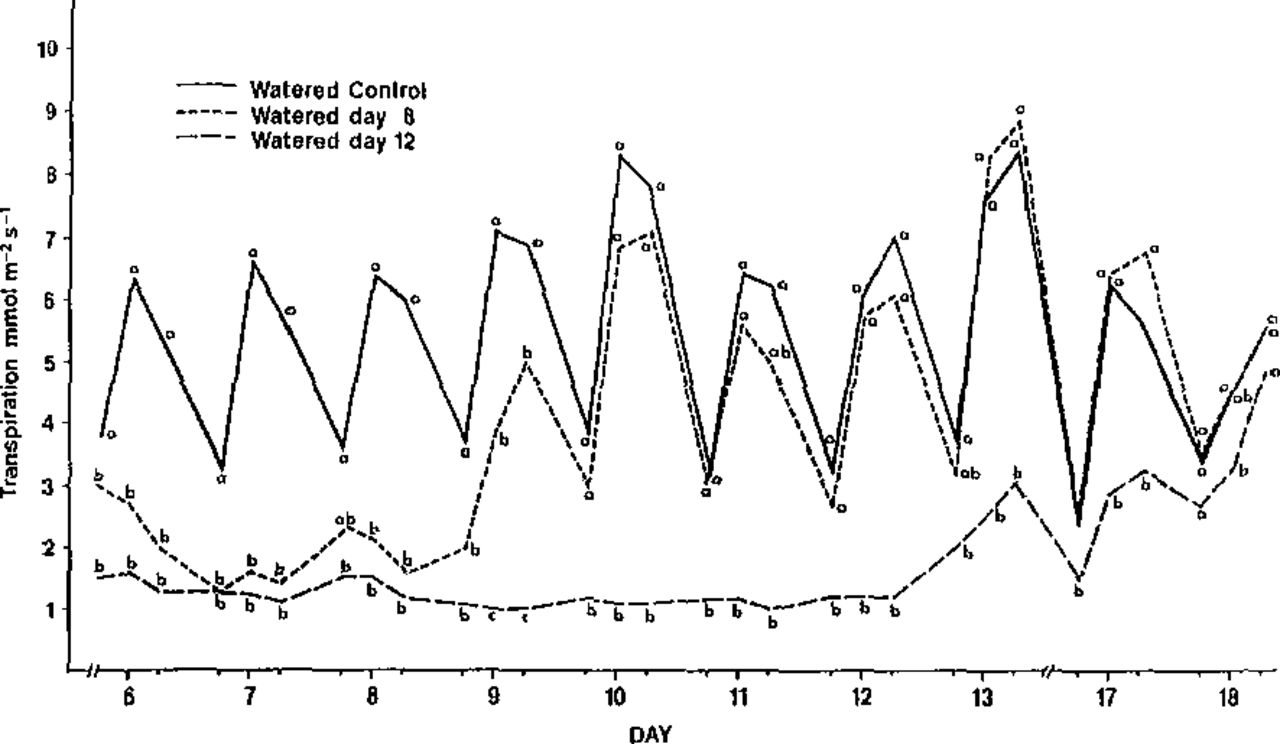
Transpiration of dogwoods watered day 8, dogwoods watered day 12 and watered control in Experiment 1; measured daily at 7:00 a.m., 11:00 a.m., and 3:00 p.m. beginning 12 June, 1984. Mean separation is by date using Duncan’s multiple range test, 5% level.

Stomatal conductance of dogwoods watered day 8, dogwoods watered day 12 and watered control in Experiment 1; measured daily at 7:00 a.m., 11:00 a.m., and 3:00 p.m. beginning 12 June, 1984. Mean separation is by date using Duncan’s multiple range test, 5% level.
Shoot water potential of plants not watered until day 8 decreased from −15 bars at 3:00 p.m., day 6, to −24 bars at 3:00 p.m., day 8 (Fig. 1). This represented more than 2X the shoot water potential of control plants (−10.8 bars). After watering at 3:00 p.m., day 8, the shoot water potential of plants not watered until day 8 recovered to the range of the watered control within 16 hr. Stomatal conductance, however, fell as low as 43.5 mmol m−2s−1 on day 8 and required 40 hr to recover (Fig. 3). Similarly, the transpiration rate of these plants which ranged as low as 1.3 mmol m−2s−1 on day 7 required 40 hr to recover (Fig. 2). Plants not watered until day 8 began to show incipient wilting on day 3. These plants wilted during the day but recovered by the following morning. This cycle continued until day 8 at 7:00 a.m. when they were at the permanent wilting point.
Shoot water potential of plants not watered until day 12 ranged from -18.0 bars to -51.3 bars (Fig. 1). After watering on day 12, shoot water potential recovered statistically to the control levels within 24 hr. The transpiration rate required up to 136 hr to return to control levels after watering (Fig. 2). Stomatal conductance returned 112 hours (Fig. 3). About 30% of the plants not watered until day 12 began to show leaf curl and some indications of marginal and tip necrosis. After watering on day 12, the foliage appeared normal by the following morning except for the necrotic areas. Even though the plants not watered until day 12 appeared to recover overnight, shoot water potential, stomatal conductance and transpiration levels took longer to return to control levels. This experiment shows that longer periods of moisture stress resulted in longer recovery periods.
Experiment 2
Suppression of growth was seen with even short periods of moisture stress. By day 15 of experiment 2, plants subjected to 5 days without water had an average shoot length of 3.9 cm, which was about half the average shoot length of control plants (Table 1). This agrees with other data that show that moisture stress suppresses shoot growth. Plants not watered until day 5 and then watered as needed had similar shoot length compared to the watered control plants by day 60. Shoot growth rate was greater from day 30 to day 120 in plants that had been mildly stressed than with the control plants. This indicates that plants undergoing mild drought stress early in the growing season compensate for initially suppressed growth by more rapid growth rate later. A short period of moisture stress did not inhibit total shoot growth over the growing season.
Longer periods of stress suppressed shoot growth further. After initial rewatering and subsequent watering, plants not watered until day 22 recovered at a slower growth rate than mildly stressed plants. By day 60, shoot length of plants not watered until day 22 was 4.7 cm, about one-third the length of the control and less than half that of plants watered day 5. However, the shoot growth of plants watered day 22 increased 3.0 cm from day 60 to 120 (2 Sept. to 4 Nov.), compared to an increase in growth of 1.0 cm in plants watered day 5 and 0 cm of the control plants. This indicates that the more severe the moisture stress, the more rapid the growth later in the season, assuming adequate moisture is provided. Yet these plants never attained growth comparable to growth of control plants and plants watered after 5 days. Thus, longer periods of drought can significantly inhibit total shoot growth over the course of a growing season, emphasizing the importance of irrigation to prevent extended periods of moisture stress.
Effects of watering stressed plants once and then restressing them were similar to uninterrupted stress. Plants watered day 5, restressed and then watered regularly from day 18 followed a similar trend to plants watered day 10. Shoot growth rate was initially suppressed by moisture stress, then began to increase slowly after watering, and increased more rapidly later in the season compared to control plants. Plants watered day 10, restressed and rewatered day 18 were similar to plants not watered day 18 and plants watered day 22 appears to indicate that a single watering during prolonged moisture stress does little to reduce the inhibitory effect of the stress on shoot growth. Both in the nursery and the home landscape, a single watering during an extended drought may be of little benefit; repeated watering is necessary during these periods.
Shoot measurements were discontinued on 4 November, 1984 (day 120) and resumed 31 March, 1985 (day 270). Spring growth flush was similar among treatments with the exception of plants watered day 22 and plants watered day 38. Plants not receiving water until day 22 had a slower growth rate in this first growth flush, illustrating the long-term effects of drought stress from one growing season to the next.
In summary, this study affirms that even short periods of moisture stress reduce stomatal conductance, transpiration, shoot water potential, and inhibit shoot growth, with effects intensified with prolonged stress. This also emphasizes the importance of regular and thorough watering during drought stress versus occasional watering of extablished trees in the landscape and trees in production nurseries. These data also show that drought stress occurring early in the season may stimulate growth later in the season as adequate moisture becomes available. We speculate that trees responding with a greater rate of shoot growth late in the season may be more susceptible to winter injury than nonstressed plants. Also, with plants severely stressed, growth the following year is suppressed, even with adequate moisture.
- © 1987, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.