
Litter from urban trees is a widespread problem, varying in magnitude among trees of many species. Vet, it is often overlooked when a city chooses trees for use along its streets. The city may plant and maintain the trees, but litter is largely a problem for householders who must endure it on their lawns and on the sidewalk and street in front of their homes. Their plight begs for attention. This paper describes typical problems of fruit litter and suggests some possible remedial strategies.
Problems
American sweetgum (Liquidambar styraciflua) is among the most extensively planted trees along urban streets in California (Barker, 1983). Numerous desirable traits account for its popularity, yet its fruit is a vexing litter problem. These dark gray, spiny balls, slightly smaller than golf balls, drop from a tree intermittently throughout the year. They are tough and leathery and disintegrate very slowly. Tall grass may conceal them but a reel lawnmower stops abruptly when one catches between blade and bar. Sometimes they are painted and used as ornaments, but this use only dents the supply. Overall, sweetgum fruit lacks redeeming qualities.
Lavalle hawthorn (Crataegus × lavallei) frequently is recommended for urban streets, and many of its traits justify the recommendation. It has attractive, dark green, leathery leaves; masses of white flowers each spring; and spectacular, brick-red fruit. Moreover, the comparatively small size of this species makes it ideal for narrow spaces along streets where its roots should not be particularly destructive to sidewalks or curbs.
Lavalle hawthorn was planted some 25 years ago in the 2½-foot-wide treelawns along several streets in the Sheffield Village district of Oakland, California. Today these trees are about 25 feet tall, probably their ultimate height. They give the streets a sedate charm (Fig. 1). Here, Lavalle hawthorn does, indeed, “fit the space.” Despite such narrow treelawns, only occasionally is there variation in surface texture or color of the sidewalks along these streets that would suggest segment replacements due to root damage.

Lavalle hawthorn trees, approximately 25 years old, line each side of Covington Street in the Sheffield Village district of Oakland, California
Yet the olive-size fruit that falls from these trees onto sidewalks creates a serious problem. People unintentionally walk on it at the risk of slipping and of staining carpets when tracking the crushed pulp indoors. It is not surprising that many residents of Sheffield Village intensely dislike Lavalle hawthorn and want the city to replace these trees with some that have no fruit.
In Davis, California, a row of six 25-foot-tall, full-grown purpleleaf plums (Prunus cerasifera) provided desirable and much appreciated shade on the west side of a residence. Nevertheless, they were recently cut down because their fruit, an intolerable nuisance, annually littered the concrete walkway that connected the front and rear yards.
Fleshy fruit usually is messy, but other types of fruit also can be annoying. Examples are pods of carob (Ceratonia siliqua), honeylocust (Gleditsia triacanthos), black locust (Robinia pseudoacacia), and Japanese pagoda-tree (Sophora japonica); ball-like fruit of planetree (Platanus × acerifolia) and sweetgum; and miscellaneous-type fruit on such tree species as carrotwood (Cupaniopsis anacardioides), magnolia (Magnolia spp.), oak (Quercus spp.), and Chinese pistache (Pistacia chinensis).
Remedies for fruit litter are not very satisfactory. Sometimes municipal tree crews remove some of the branches of a street tree loaded with annoying fruit, temporarily reducing the potential for fruit litter. When in flower, trees can be sprayed with a growth hormone to prevent fruit set. But this treatment is neither permanent nor foolproof. Moreover, people increasingly object to tree spraying in urban areas, whatever the purpose. Tree removal is another possibility but only occasionally has fruit litter alone resulted in such drastic action. Householders who have a littering tree, therefore, usually have little choice but to be ambivalent: accept the tree for its benefits but hate it for its litter.
Fruitless Selections
If little can be done to prevent or diminish the problem of fruit litter of existing trees, possibly the best solution, in the long run, is to use nonfruiting trees in newly developed areas or when existing trees are replaced.
Named cultivars considered to be non-fruiting are available for a few species, notably ash (Fraxinus spp.), honeylocust (Gieditsia triacanthos), and olive (Olea europaea). But for most species, non-fruiting individuals have yet to be identified and cloned for distribution by the nursery industry.
Through regular monitoring of a population of trees of a species, the level of variability in fruit yield among them can be determined and nonfruiting or, as an alternative, scant-fruiting individuals identified. To illustrate, 50 European hornbeam (Carpinus betulus) trees are located along a street in Davis, California. Planted as 5-gallon stock in 1966 and 1967, they vary in height from 20 to 45 feet. I rated them for fruit yield for three consecutive years, beginning in 1984 (unpublished data). Rating was on a scale of 1 (heavy fruiting), 4, 6, 8, and 10 (no fruit).
On seven of these trees there was no fruit any year, making them promising candidates for selection for fruitlessness for reasons mentioned later. (The one tree of this group that I checked two years for flowers had none.) Three other trees lacked fruit two years and had only scant fruit (8 rating) a third year. At the other extreme, on 15 trees, there was heavy to moderately heavy fruiting (4 rating) all three years. With the remaining 25 trees, or one half the population, there was moderate fruiting. Among the three years, the fruit yield of each of these 25 trees was fairly consistent except for a few of them that each had yields at or near opposite ends of the rating scale.
European hornbeam fruit consists of a nutlet, slightly less than pea size, attached to a 3-lobed bract that is about one half the length of a typical leaf of this species. Both the nutlet and the bract dry and turn straw color before falling in August or September. Although this fruit, which resembles leaves, may be a visual nuisance when it collects on lawns, its primary nuisance, like the fruit of many other tree species, is germination of the seeds that fall into flower beds and other bare-soil areas and subsequent growth of unwanted seedlings. For this reason, a non-fruiting cultivar of European hornbeam, which the foregoing data suggest could be developed, should be distinctly superior for urban use.
Besides the above mentioned non-fruiting hornbeam trees that may also be non-flowering, a unique sweetgum, identified by Santamour and McArdle (1984) as (Liquidambar styraciflua forma rotundiloba), possibly is non-flowering. This tree was found about 1930 in a natural stand of second growth mixed hardwoods near Cameron, North Carolina, about midway between Sanford and Southern Pines (pers. comm. Eloise Knight). It differs from other trees of the species by having rounded instead of pointed leaf lobes. Neither the original tree nor any of its self-rooted propagules are known to have flowered yet they each match other trees of the species in having a distinct central leader (pers. comm. Herbert Hechen-bleichner, Lionel Melvin, Herbert P. Smith).
The biological basis for fruitlessness of trees differs within and among species. If trees never flower, they obviously never fruit. Other bases for fruitlessness are flower sterility and flower type. Assessment of the potential for non-fruiting individuals of a tree species is simplified by sorting an array of species into categories according to their reproductive morphology. Species in Category A of Table 1, for example, have bisexual flowers. Existence of non-fruiting individuals in these species most likely would be due either to lack of flowers or flower sterility.
Tree taxa classified according to their generally recognized reproductive morphology.
Trees with sterile flowers or few if any flowers would just as likely exist in the remaining species of Table 1 (Categories B and C). Even so, with these species, it may be expedient in developing non-fruiting individuals to select trees with only male flowers. One of the easiest ways, for example, to obtain a non-fruiting tree is to propagate a male individual of a dioecious species vegetatively. Ginkgo {Gingko biloba) is dioecious. Autumn Gold ginkgo, a cultivar of this species, is male. Chinese pistache is another dioecious species. Increasingly popular for urban uses, its fruit resembles small peas and creates sufficient litter to justify clonal propagation of male trees.
Kentucky coffeetree (Gymnocladus dioicus) likewise is dioecious. Female trees have exceptionally large woody pods, ranging from 4 to 6 inches long and 1 to 2 inches wide. These pods remain on the tree long after they have ripened and opened. A substance that leaches from the opened pods during rains is highly corrosive to automobile finishes. On the other hand, male trees of this species should be ideal for urban use because the species generally is free of pests and widely adaptable. Also desirable is the tree’s sparse branch structure that blocks little solar radiation in winter.
Trees having only male flowers are not limited to dioecious species. A few individuals of either a monoecious or a polygamous species may be unisexual. California buckeye (Aesculus californica), for example, is considered polygamous, with both male and bisexual flowers on the same tree. Numerous traits of this buckeye make it promising for urban use. Intermediate in size, structurally strong, drought tolerant, and early to defoliate, it is compatible around homes utilizing solar energy. However, its fruit, which is slightly larger than a golf ball, can be dreaded litter.
Benseler (1968) surveyed numerous natural populations of California buckeye throughout Northern California and found individuals with only male flowers, although none was specifically identified. Encouraged by this finding, in summer 1982, I searched natural populations of California buckeye in Solano County, California, and found several individuals that had no fruit. At flowering time the following spring, 1983, I revisited these non-fruiting trees and found that most of them had bisexual as well as male flowers, indicating their potential to fruit in the future. Two individuals, however, indeed had only male flowers in 1 983 and in each year since. These two trees are candidates for introduction as non-fruiting cultivars of California buckeye.
American sweetgum typically is monoecious, with both male and female flowers on the same tree. Might some individuals have only male flowers? Inflorescences of tiny flowers open along with the leaves in early spring. Male flowers in small sessile and subsessile heads form a terminal raceme (Fig. 2). Small stalked heads near the base of the inflorescence typically comprise female flowers, although some of these also have only male flowers. Shortly after pollen release the male portion of the inflorescence abscises and any stalked heads of female flowers remain to develop into fruit. Although most inflorescences have one or two stalked heads of female flowers a few of them have none, suggesting the possibility of finding a tree that has only inflorescences with heads of male flowers exclusively.
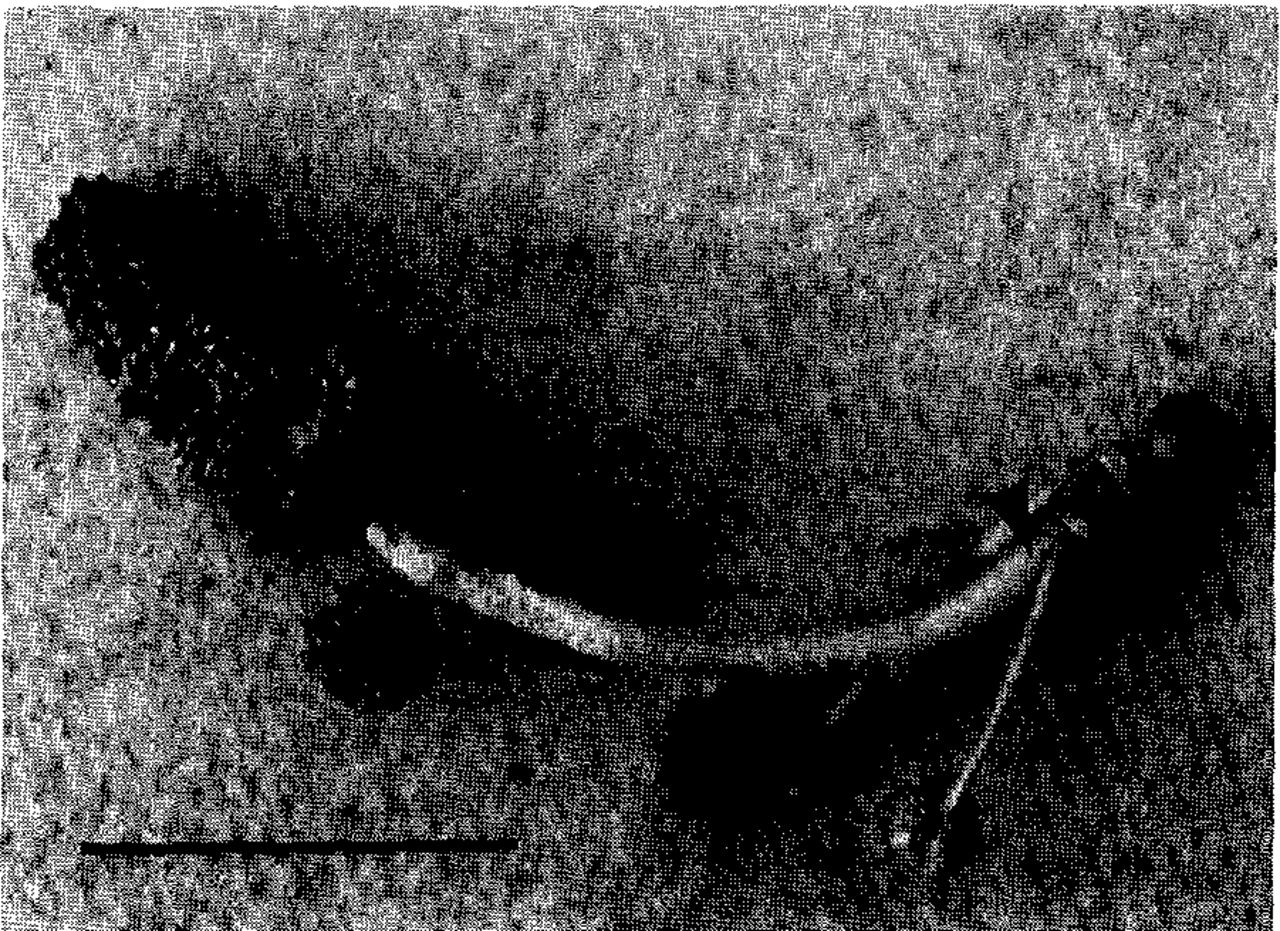
An American sweetgum inflorescence, comprising a terminal raceme of sessile and subsessile heads of male flowers and one stalked head of female flowers at the base. The arrow shows where the raceme portion of the inflorescence abscises following release of pollen from the male flowers. Scale: bar = 1 inch.
Any sweetgum trees that are non-fruiting because they have no flowers or else have only male flowers should be prime candidates for selection and followup evaluation for possible introduction as fruitless cultivars.
In the selection of non-fruiting trees of any species appearing to have only male flowers, it is prudent to be cautious. A few trees of a dioecious species may be monoecious or polygamous, as reported for ginkgo by Miyoshi (1931), or dioecy may prevail in a few trees of monoecious or polygamous species. Or the flowering pattern of an individual tree may differ from year to year. Canyon maple (Acer grandidentatum)), for example, typically has male and functionally female flowers on the same tree and a few trees apparently have only male flowers. Randomly selected trees of this species in natural stands in Utah were studied for two consecutive flowering years (Barker et al., 1982). After a severe drought that began in summer and continued through winter an unusually high frequency of the trees had only male flowers. However, many of the exclusively male trees had old fruit stalks; proof that they had had female flowers in previous years. Similarly, production of fruit on asexually propagated, supposedly “male” cultivars of ginkgo has been reported (Santamour et al., 1983). And in a plantation of more than 300 seed-propagated ginkgo trees, each 35 years or older, a few trees produced fruit some years, although apparently an insignificant amount, and only male flowers other years.
Why such sexual flexibility? Probably because sex expression of a plant not only is genetically controlled, but may also be affected by weather patterns and the physical environment, with stressful events triggering a shift in some plants’ normal sexual phenotype (Barker and Freeman, 1979; McArthur and Freeman, 1982).
Floral primordia of plant species that typically have unisexual flowers of a particular gender evidently have the potential to develop as either bisexual or opposite-gender flowers. This has been demonstrated in California buckeye. Inflorescences of this species comprise 100 or more flowers on a shaft or axis 8 to 12 inches in length. Only a few flowers near the tip of the axis have fully developed pistils, are bisexual, and capable of producing fruit. The other flowers in an inflorescence typically are male with only rudimentary pistils. During inflorenscence development, elongation of the axis of the inflorescence precedes opening of the individual flowers. To study gender conversion possibilities of the male flowers, I cut the axes of inflorescences back to about one half their original length. This was done when the flowers of the inflorescences were still in bud stage. Typically, the remaining segment of each shortened axis would have only male flowers. Yet, by the time the flowers opened, a few of the terminal ones on many of these shortened axes had fully developed pistils, from which fruit subsequently developed (unpublished data).
Ultimate gender of a flower—bisexual, male, or female—may be regulated by the relative amounts of different hormones present in various tissues of the flower during its development (Torrey, 1976). The concentration of at least some of these hormones depends partly on the environment. Drought, as an example, suppresses biosynthesis of cytokinin (Itai and Vaadia, 1970), a hormone that promotes development of female tissue of the normally male Sultan grape (V/tis vinifera) (Negi and Olmo, 1966) and of normally male flowers of spiderflower (Cleome iberidella) (Jong and Bruinsma, 1974).
Although proof is lacking, cytokinins may affect development of female tissue in the flowers of tree species with reproductive systems similar to the above-mentioned grapes and spiderflower, including various species of maple (Acer) (Jong, 1976), buckeye and horsechestnut (Aesculus) (Benseler, 1975), carrotwood (Cupaniopsis) (Bawa, 1977), persimmon (Diospyros) (Hodgson, 1939), and olive (Olea) (Brooks, 1948; Hartman and Panetsos, 1961). If true, then trees of such species that lacked female flowers in a dry environment may have both female flowers and fruit in a more favorable environment, as reported for canyon maple (Barker et al., 1982).
Besides drought, other environmental factors that inhibit plant growth may also affect sex expression. One of these is low light intensity, which suppresses carbohydrate synthesis. Red buckeye (Aesculus pavia) plants growing on woodland sites under low light intensities were found to have bisexual flowers less often than did plants in open fields (Bertin, 1982). Similarly, in pawpaw (Asimina triloba), flowers on shaded stems had proportionately fewer female parts than those on sunlit stems (Willson and Schemske, 1980).
Finding Non-fruiting Trees
Caution notwithstanding, opportunities abound for monitoring populations of trees in search of non-fruiting individuals. This could be done for each of various species in many parts of the country, in both natural stands in rural areas and in planted populations, as along streets in urban areas.
Selecting non-fruiting individuals, getting clones of these produced, and planting them instead of fruit-bearing trees of the same species may be the best long-term solution to the problem of fruit litter of urban trees.
Footnotes
↵1 Updated version of a paper presented at the annual conference of the Western Chapter, International Society of Arboriculture, Palo Alto, California, May 19-21, 1984.
- Copyright © 1986, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.