
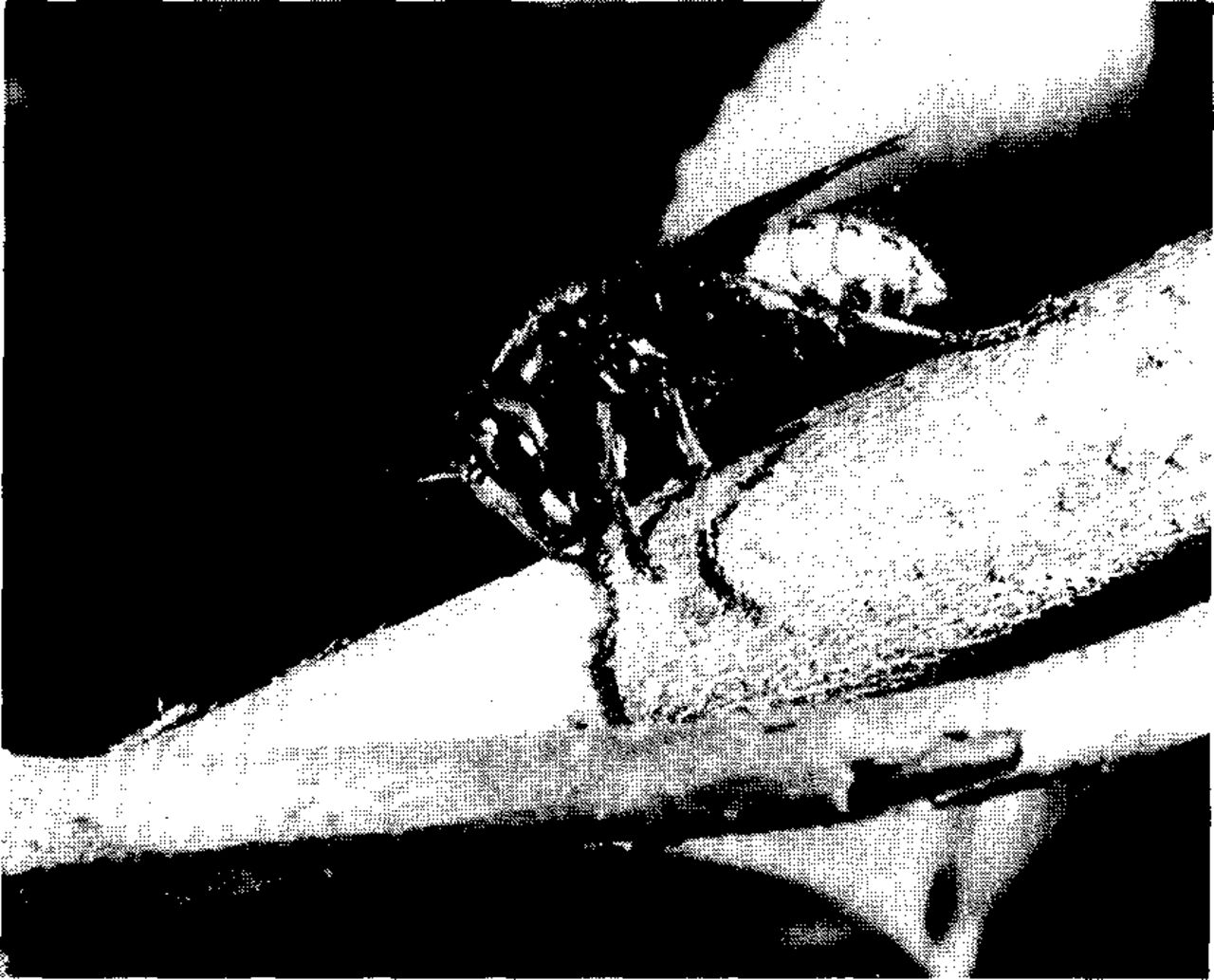
On September 6, 1984, while surveying our provenance plantings of green ash (Fraxinus pennsylvanica) for seed production, we encountered a curious situation. Large wasps were “attacking” various branches in several trees and, working individually or in small groups (Fig. 1), appeared to be stripping the bark from these branches. Several of these attack sites already had several cm2 of bark removed on that date. This event prompted us to conduct a survey of damage in all of our ash plantings during the dormant season of 1984-85. Likewise, in the spring of 1985, while bagging branches of various birches for controlled pollination, we saw similar damage (presumably from 1984) in many trees; another damage survey was made in our birch plots.

European hornet worker removing bark of Chinese ash.
The insect responsible for the damage was the European hornet, Vespa crabro. We assumed that, since we were now aware of the problem, we would be able to monitor hornet activity closely in 1985. Unfortunately, there was no further activity in the plantings. However, we were able to make some significant observations that year on bark stripping behavior by hornets at another location that were working on a Chinese ash, grown from seed received in 1929 as F. chinensis var. rhynchophylla (Hance) Hemsl. (= F. rhynchophylla Hance).
The Insect
According to information summarized by Shaw and Weidhaas in 1956 (15), the European hornet was introduced into the United States between 1840 and 1860. Its present distribution ranges along the eastern seaboard from New England south into northern Georgia and Alabama and westward through Tennessee to the Mississippi River north to Michigan and southern Ontario (1). Scattered populations may occur south to the Gulf of Mexico and west through Illinois, Iowa, Wisconsin, and Minnesota.
The adults of the European hornet resemble large yellowjackets and are easily distinguished from the common eastern yellowjacket {Vespula maculifrons) and the so-called baldfaced “hornet” {Dolichovespula maculata) (Fig. 2). Nests are composed largely of decayed wood and located in cavities in trees, buildings, or other sites above ground where the fragile brown or tan nest material is somewhat protected from the environment.

Workers (below) and queens (above) of, left to right: European hornet, baldfaced hornet, and eastern yellowjacket. Pale markings on baldfaced hornet are white; yellow on others. European hornet worker Is about 25 mm long.
The first report of tree damage by this insect in the United States was made by Thomas Meehan, a Philadelphia nurseryman, in 1878 (10). (Although Meehan was under the impression he was observing D. maculata, it is clear from his description that the insects were indeed V. crabro.) He reported that young branches of European ash (Fraxinus excelsior) and common lilac (Syringa vulgaris) had been stripped of bark by the hornet. Cory (4) provided additional information on damage to lilac, which seemed to be “the preferred woody plant” for hornets in Maryland. Lilac was also the primary host noted by Shaw and Weidhaas (15), who also reported damage on syringa (Philadelphus sp.) and cited earlier reports on ash, birch, dahlia, and horsechestnut. Other references mention the hornets damaging alder, elm, dogwood, rhododendron, boxwood, and fringetree (Chionanthus virginicus) (2,7,8). None of these observations recorded any quantitative data relating to differences in damage among genera, species, or cultivars.
The Trees
Our ash and birch plantings provided an excellent opportunity to make observations on hornet attack on trees of different species and geographic origins. All of the trees were located in the National Arboretum‘s test area in Beltsville, Maryland, at approximately 39° N. Lat. and with an average January temperature of 33.5°F (ca. 1°C).
Most of the ash trees were green ash (Fraxinus pennsylvanica) and were contained in 4 major plantings. Two of these (FP-1 and FP-2) were replicates of a provenance test that included trees from 41 native origins in 19 states and 5 Canadian Provinces throughout the species‘ range. The seedlings had been planted as 2-0 stock in 1978, in 4-tree row plots at a spacing of 10 feet between trees and rows. Inasmuch as most plots contained seedlings from more than one tree in .each provenance, there was a wide range of plant genetic diversity available to the insect. Two seedlots of white ash (F. americana) from Ohio and Illinois were also included in these replicates. The 2 replicates were located on quite different sites, but were only 100 yards apart.
The third planting of green ash (FP-3) consisted of seedlings from individual mother trees from 19 geographic origins, and was planted in 1980 with 2-0 stock in unreplicated 4-or 5-tree row plots. The fourth planting (FP-4) was made in 1979 and contained progeny from a single native tree on the National Arboretum grounds in Washington, D.C. These seedlings were originally thought to represent a number of interspecific hybrid combinations from semi-controlled pollinations, but all had proved to be pure green ash.
All of the ash plantings were no more than 200 yards distant from one another. In addition, there was a young planting of F. sieboldiana located adjacent to FP-2 on the side closer to FP-1. These trees were only 4 to 6 feet in height at the time of the survey.
The birch trees represented the remnants of test plots planted in 1973 to evaluate various species and geographic origins of the 5 major white-barked birches: Betula papyrifera and B. populifolia from the United States; fl. pendula and fl. pubescens from Europe; and fl. platyphylla var. japonica from Japan. The early growth of these trees and their relative susceptibility to bronze birch borer have been reported previously (13, 14). Because of mortality or stem dieback that resulted from borer attack, many trees were either dead or represented only by a major trunk 6 feet to 10 feet in height derived from a basal or lateral sprout. However, even though the original randomization of the planting design had been destroyed, there remained a sufficient intermixture of species and origins to allow the hornets considerable freedom of choice in host plants. The birch planting was located roughly 300 yards from the ash plantings.
The Damage
In surveying the damage in plantings of both genera, individual trees were classified only as “attacked” or “not attacked,” with no attempt to quantify the number of attack sites or the area of bark removed. Such attention to detail may have had some importance, but the removal of some damaged branches and even entire trees for other research studies in 1985 precluded accurate comparisons at a later date.
In neither ash nor birch did the hornets strip bark from young branches only one season old. Rather, they seemed to favor branch sections 3 to 5 years old with thicker bark. Branch diameter at this age generally ranged from 1.5 cm to 3.0 cm at the time of attack. We did find evidence of attempts to strip bark from larger and older branches, but the hornets apparently had abandoned these sites before they had chewed down to the cambium and removed any bark.
The amount of bark removed at any one site on the ash trees varied from small patches only about 1 cm2 to long irregular patches that girdled the branch (Fig. 3). When ash branches were not girdled, vigorous callus growth was initiated the following season, sometimes completely covering the bare wood that had been previously exposed by bark removal. Determination of the cause of the “injury” after callusing would be difficult in some instances (Fig. 3). However, where the bark patches removed were relatively small and regular, characteristic radiating lines in the young callus, indicating growth from all 4 sides of the wound, at least would suggest a physical loss of bark rather than a canker disease, borer site, or bird damage.
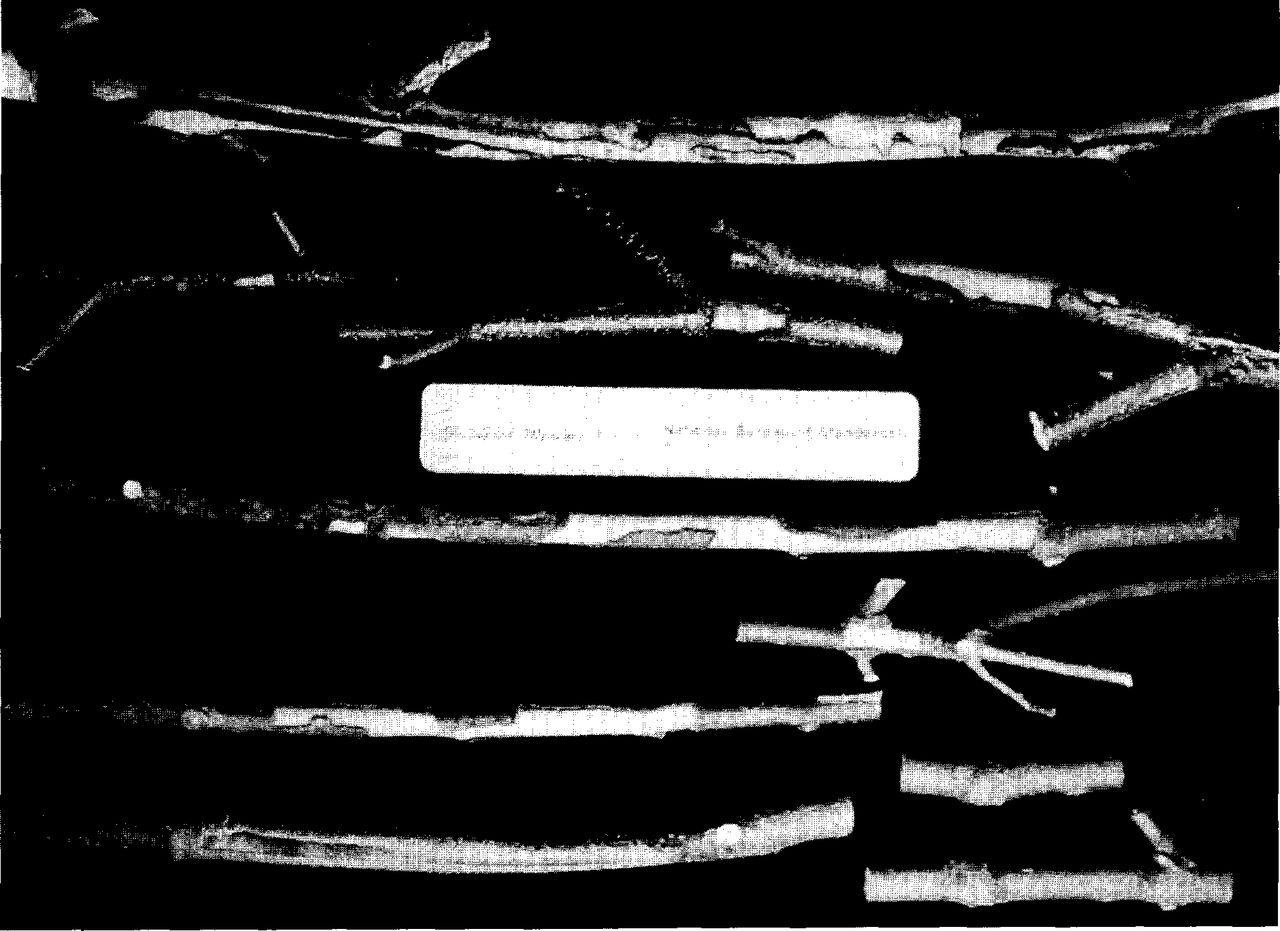
European hornet damage on (above ruler) Japanese white birch and (below ruler) green ash. The upper branch of birch was girdled and was killed. Other branches were not girdled but little callus formed. Upper branch of ash was girdled and middle 2 branches were not. These branches were removed during the dormant season following attack. Lower 3 branches, not girdled, were left on the trees for 1 growing season and produced much callus. Two branches at lower right were typical of reduced damage on trees of Washington, D.C. origin. Ruler is 15 cm long.
The hornet damage to birch branches seemed to be more extensive and destructive than on ash. With few exceptions, the branches were girdled, callusing was minimal, and the distal portion of the branch died (Fig. 3). Furthermore, there were fewer attack sites per tree on birch than on ash, but this may have been more a reflection of growth habit than generic classification.
Obviously, European hornet damage is not a life-threatening scourge of even those genera most favored by its attention. However, there may be instances in both nursery and landscape plantings when mass attacks could cause significant damage by reducing the salability of nursery stock or altering the crown growth pattern of planted specimens. It may even be that the callus growth areas resulting from bark removal would be ideal sites for egg-laying of borers. We do not want to overstress the destructiveness of the European hornet, but we do want arborists and nurserymen to be aware of and recognize the injuries caused by this insect.
Results and Discussion
When compiling the damage data from the 1984 survey of the green ash provenance test, it was noted that trees from more northern, colder areas seemed to be less likely damaged than those originating in warmer climates. In FP-1, for instance, none of the trees (85) from 22 provenances was damaged. In 20 of those provenances, the average January temperature was below 0°C (32°F). Conversely, all of the trees (32) of 9 provenances were damaged and in 7 of these the average January temperature was above 0 °C (32 °F). Therefore, the data in Table 1 are presented according to that classification. There may be little difference in the climates of areas with average January temperatures of + 1°C and -1°C and, consequently, few physiological differences in the vegetation that has become adapted to each climate. Still, 0°C appears to be a significant, as well as convenient, dividing point between “southern” and “northern” sources. It is obvious from the table that green ash trees from “northern” sources were less likely to be attacked by hornets than those from “southern” habitats. The mother tree of the progeny in FP-4 was native to a “southern” area about 1 0 miles from the planting site, which is itself a “southern” site. Of the white ash trees included in the green ash plantings, 2/8 of those from a “northern” source in Ohio were attacked, as were 3/8 from a “southern” Illinois source.
Numbers and percentages of green ash (Fraxinus pennsylvanica) trees of different origins damaged by European hornet—1984
A reasonable explanation of the observed preference of the hornet for trees of “southern” origin could be based on their lengthened period of cambial activity and bark “peelability” as compared to trees of “northern” lineage. In fact, we have shown, by monitoring these selfsame trees during the 1979 growing season, that the time of cessation of cambial activity, as measured by cambial electrical resistance, varied inversely with the average January temperature of the seed source (12). It may logically be assumed that “peelability” or “juiciness” of the bark would follow a similar pattern. When making the reciprocal patch grafts reported in that paper (12), we found that on August 8, 1979, the bark of trees from the most northern origin (Manitoba) was difficult to remove, and it is likely that loss of physical “peelability” precedes the electrically detectable cessation of cambial activity by weeks or months. Thus hornet attacks will be more numerous, and the bark area of the damage more extensive in trees from more southern origins (see Fig. 3).
The birch data in Table 2 are in general agreement with this hypothesis, although now the picture is confounded by possible species differences. Nevertheless, fewer trees from northern latitude sources were attacked.
Damage by European hornets to various species and geographic sources of white-barked birches (Betula)—1984
As noted above, hornets were observed stripping bark from F. rhynchophylla in Glenn Dale, Maryland, in 1985. No damage was observed in 1984 on the young trees of F. sieboldiana located near the green ash plots. Without complete documentation of origin, no conclusions can be drawn regarding the reasons for this disparity.
Some hornet damage from 1984 was also found (in 1985) on ash trees on the grounds of the National Arboretum. Two trees of each of 2 F. americana cultivars had been planted, along with trees of other species, in an isolated test area in 1980. There was no damage to trees of ‘Autumn Applause‘ whereas both trees of ‘Champaign County‘ exhibited numerous callused-over attack sites.
To our knowledge, bark stripping has not been reported for any other species of social wasp besides Vespa crabro. As noted by Shaw and Weidhaas (15), there has long been uncertainty in the literature as to why European hornets exhibit this behavior. Some authors have claimed the bark is gathered for use as nest material (e.g., 3, 7, 16), while others acknowledge this as a possibility but emphasize the aspect of sap collection (e.g., 1, 4, 5).
Our observations of hornets working on Chinese ash in 1985 and common lilac in 1978 clearly demonstrated that the insects were not collecting fiber. Despite extended bouts of vigorous gnawing, the bark removal was an extremely gradual, almost imperceptible process. Rather than being efficiently scraped away in strips over a period of several minutes and retained as a ball of soft pulp (the way nest material is gathered), the bark was laboriously worn away in tiny bits which apparently dropped to the ground. Some individuals worked at the same spot for several hours with little or no visible progress. Meehan‘s (10) reference to hornets carrying “strips of bark away in their mouths” is therefore puzzling, not only because social wasps do not transport fiber in this manner, but also because the hornets seemed incapable of removing the bark in strips.
It was equally clear that considerable sap collection was taking place. Bark gnawing alternated with usually longer periods of lapping at the freshly exposed cambium. Sap flow from wounds on the Chinese ash was copious at times. In many cases the stripped patches appeared dry, although the abundance of files, ants, yellowjackets (Dolichovespula and Vespula spp.) and paper wasps (Polistes spp.) at these areas attested to the nutrients present. The feeding hornets repeatedly lunged at these intruders to drive them off, occasionally taking a yellowjacket or fly as prey.
In fact, the attraction of Vespa crabro to these sources of sap and the effort expended to secure it goes far beyond any other known type of social wasp foraging behavior. Considering the amount of damage we observed in the plantings coupled with how slowly the bark was actually removed, an enormous number of wasp-hours must be devoted to this task. At least 140 trees in the same area displayed some damage and up to ca. 80 hornets were counted per tree. The propensity for nocturnal foraging by V. crabro must also be taken into account. Observations in late September revealed as many hornets on the Chinese ash at night as were present during the day. With temperatures too cold for flight, the insects slowly and feebly exhibited chewing and lapping behavior or just remained motionless with mouthparts on the cambium.
Inasmuch as the European hornet is not considered an abundant species in this country, and typical mature nests contain less than 500 workers (1), bark stripping must therefore constitute a major fraction of the time and energy budgets of at least some colonies in late summer and fall. An important question is whether this behavior is adaptive for the insects, and thus represents mining for essential nutrients which cannot be obtained elsewhere, or whether it is actually a detrimental preoccupation with highly attractive but non-essential compounds and represents a drain on colony resources. Vespa spp. characteristically feed on naturally oozing sap, and this is considered to be the most important carbohydrate source for the 5 species occurring in Japan, including V. crabro (9). Interestingly, bark stripping to obtain sap has not been recorded in any region other than Europe and North America.
Another major question is why ash and birch are preferred sap sources. As far as we could determine from the literature (6), there is little similarity in bark or phloem (sap) chemicals in these 2 genera. In fact, there are great differences in the presence of coumarin compounds in various ash species: absent in F. americana and F. Pennsylvania; strongly present in F. excelsior, F. rhynchophylla and F. sieboldiana (11). Oddly enough, oaks (Quercus spp.) are considered the most common source of naturally exuding “sap” for hornet species, including V. crabro (9), and is the only such source on which we have observed them in Maryland. Yet we are unaware of any records of bark stripping on oak. (A reasonable explanation for this, however, is that hornets on oak are not actually feeding on sap, but on the fermented product of bacterial wetwood infections.)
Many other kinds of trees occurred near the ash and birch plantings, but were ignored by the hornets. These included sweetgum (Liquidambar styraciflua), tuliptree (Liriodendron tulipifera), honeylocust (Gleditsia triacanthos), planetree (Platanus hybrids), and red maple (Acer rubrum). At this point, the critical factor or factors shared by all of the hornets‘ preferred sap sources remain a mystery.
Footnotes
↵1. Respectively, Research Geneticist, U.S. National Arboretum, and Research Associate, Department of Entomology, University of Maryland, College Park, MD.
- Copyright © 1986, International Society of Arboriculture. All rights reserved.
In this issue








{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.